
Phénomènes électriques en rapport avec l'influx nerveux - Ts
Classe:
Terminale
Thème:
Le tissu nerveux et ses propriétés
Introduction :
L'une des propriétés de la cellule nerveuse est la conduction de messages nerveux appelés influx nerveux.
Cette conduction est en rapport avec des phénomènes électriques au sein de la cellule nerveuse.
Ces phénomènes électriques peuvent être décelés à l'aide d'appareils spécifiques.
Mais aussi, ils peuvent être expliqués de manière ionique.
I. Principe de fonctionnement de l'oscillographie ou oscilloscope cathodique
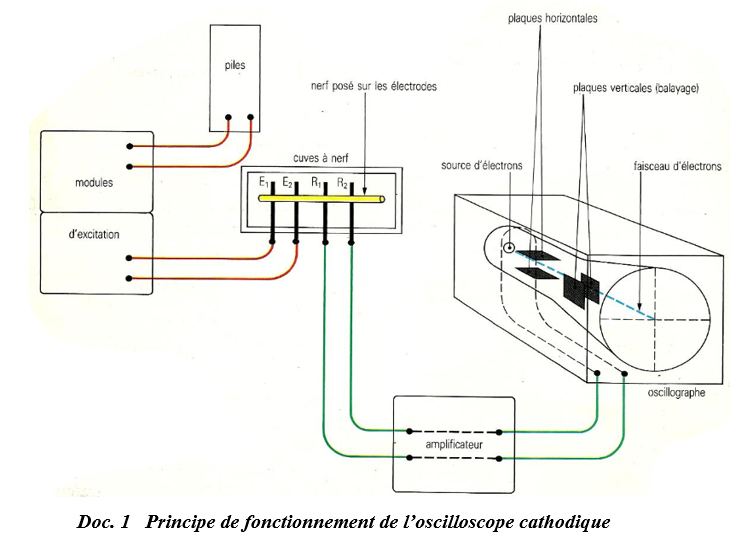
II. Mise ne évidence des phénomènes électriques
Expérience
On met en contact les faces externes et internes d'un neurone avec un galvanomètre
Résultats
On note une déviation de l'aiguille de l'appareil
Interprétation
Le mouvement de l'aiguille met en évidence des phénomènes électriques au niveau de la fibre nerveuse.
Ces phénomènes sont dus à une $ddp$ et le sens de déviation de l'aiguille montre que l'extérieur de la fibre est chargé positivement et l'intérieur négativement
Cette polarisation électrique observée au niveau de la fibre est commune à toutes les cellules vivantes.
II.1 Le potentiel de repos
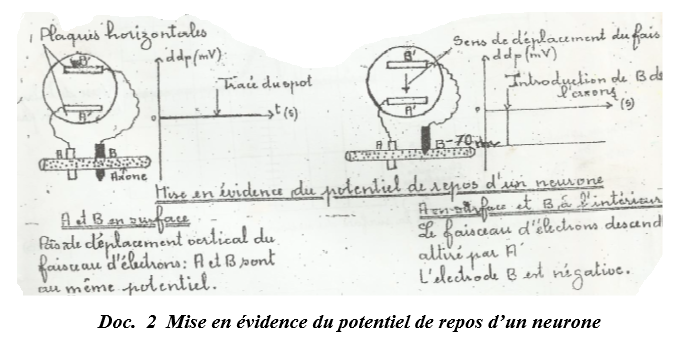
Les enregistrements se font à l'aide d'un oscillographe muni d'électrodes réceptrices et d'un stimulateur muni d'électrodes stimulatrices.
La $1^{ière}$ et la $2^{ième}$ électrode sont respectivement en relation avec la plaque horizontale inférieure et supérieure de l'appareil
$\bullet\ $ Si les deux électrodes réceptrices sont placées à la surface de l'axone, en dehors de toute stimulation, le spot décrit une ligne horizontale au niveau zéro sur l'écran de l'oscillographe.
Le résultat est le même quel que soit la position des électrodes à la surface de l'axone.
On peut dire que tous points de la surface du neurone au repos sont au même potentiel.
$\bullet\ $ Si on enfonce l'électrode $B$ à l'intérieur de l'axone, le spot dévie brusquement.
C'est la preuve qu'il existe un $ddp$ entre la surface et l'intérieur du neurone $(ddp=-70\,mV$ chez le calmar, $ddp=-60\,mV$ chez les mammifères).
Cette $ddp$ est appelée potentiel de repos ou potentiel membranaire du neurone qui par définition est le potentiel de membre en dehors de toute stimulation.
Donc on peut dire qu'au repos, le neurone est polarisé électriquement.
II.2 Potentiel d'action

Il existe deux types de potentiel d'action : le potentiel diphasique et le potentiel monophasique
II.2.1 Potentiel diphasique
Si on place les électrodes réceptrices $A$ et $B$ à la surface de l'axone et qu'on porte une stimulation d'intensité suffisante sur la fibre nerveuse, il apparait une perturbation sur l'écran de l'oscillographe.
En effet, le spot décrit une onde dissymétrique avec deux pointes appelées spikes.
Cela traduit l'existence d'une $ddp$ provoquée d'abord en $A$ puis en $B.$
Cette $ddp$ est le potentiel d'action de la fibre nerveuse.
Par définition le potentiel d'action est la $ddp$ membranaire suite à une stimulation suffisante ou la perturbation du potentiel de repos suite à une stimulation suffisante.
Interprétation : (du tracé sur l'oscillographe)
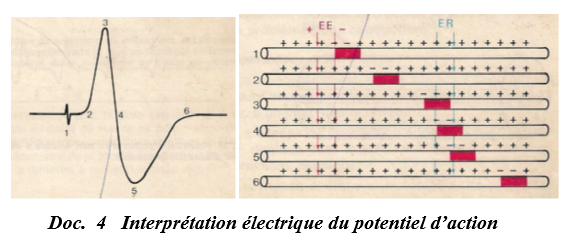
$-\ $ En $1$, c'est l'artéfact de stimulation ; il correspond au moment de la stimulation (la vitesse du courant électrique étant supérieure à celle du courant biologique ou influx-nerveux)
$-\ $ Entre $1$ et $2$, il ne se passe rien ; c'est le temps de latence qui correspond au temps que met l'influx nerveux pour arriver à la première électrode réceptrice $A.$
$-\ $ Entre $2$ et $3$, la déviation du spot vers le haut indique que l'électrode réceptrice $A$ est devenue négative, il y a alors une inversion de la polarité membranaire par rapport à l'état de repos : c'est la dépolarisation sous l'électrode réceptrice $A$
$-\ $ Entre $3$ et $4$, la descente du spot montre un rétablissement des potentiels entre $A$ et $B$ : c'est la repolarisation l'électrode réceptrice $A.$
$-\ $ Entre $4$ et $5$, la déviation du spot vers le bas montre que maintenant, c'est l'électrode $B$ qui devient négative : c'est la dépolarisation sous l'électrode réceptrice $B.$
$-\ $ Entre $5$ et $6$, la déviation du spot vers le haut explique que l'électrode $B$ tend à devenir positive : c'est le repolarisation l'électrode réceptrice $B$
$-\ $ En $6$, c'est le retour à l'état initial car l'onde à dépasser les deux électrodes réceptrices $A$ et $B.$
NB :
L'influx nerveux est une onde négative qui se propage à la surface de la fibre nerveuse et son enregistrement à l'oscillographe montre des potentiels d'actions.
Remarque
$-\ $ Si les électrodes $A$ et $B$ placées à la surface du neurone sont relativement proches l'une de l'autre, on obtient une courbe diphasique dissymétrique car la dépolarisation sous la deuxième électrode $B$ a commencée pendant que la repolarisation sous la première électrode $A$ n'est pas entièrement achevée :
On obtient une courbe d'amplitude plus faible que celle de la première partie (doc. ).
$-\ $ Si les électrodes $A$ et $B$, toujours placées à la surface de l'axone sont suffisamment éloignées l'une de l'autre, on obtient une courbe diphasique symétrique car la dépolarisation sous $B$ se déroule au moment où la repolarisation sous $A$ est entièrement achevée.
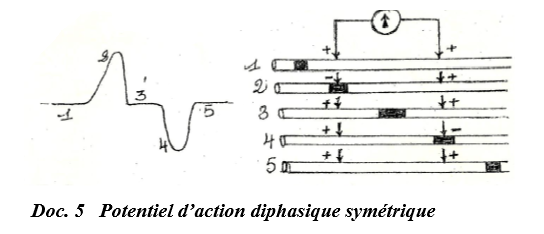
II.2.2 Potentiel monophasique
Il est obtenu soit en mettant en contact une électrode avec la surface d'une fibre et l'autre, comme électrode de référence, soit en maintenant une électrode en surface et l'autre, à l'intérieur de la fibre.
Dans le deuxième cas on notera une hyperpolarisation due à une augmentation de la $ddp$ quand l'onde arrive au niveau de la deuxième électrode réceptrice $B.$
NB :
$\bullet\ $ Si la stimulation est suffisante, on a un potentiel d'action reconnaissable par l'inversion de signe et qui se propage sur toute la fibre nerveuse.
$\bullet\ $ Si la stimulation n'est pas suffisante, on a une dépolarisation locale encore appelée dépolarisation de membrane qui ne se propage pas.
Remarque :
Notion de potentiel récepteur
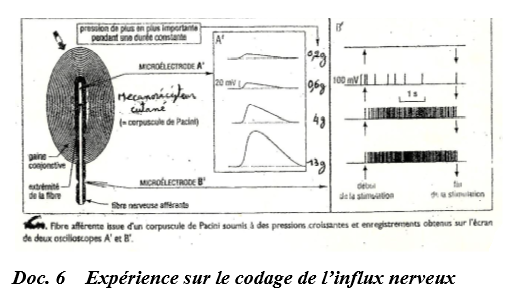
Au niveau d'un récepteur sensoriel, une fibre nerveuse répond à une stimulation par une dépolarisation locale d'amplitude graduable en fonction de l'intensité du stimulus.
Cette réponse constitue le potentiel de récepteur qui traduit l'intensité du stimulus en amplitude de potentiel de récepteur.
Ce potentiel de récepteur, au-dessus d'une valeur seuil engendre (au niveau de la fibre sensitive) un train de potentiels d'action d'amplitude constante mais de fréquence proportionnelle au potentiel récepteur.
On dit que l'information initiale codée en amplitude au niveau du récepteur est convertie, au niveau de la fibre, en message codé en modulation de fréquence.
La transformation de l'énergie du stimulus en un message de type nerveux est appelée transduction.
III. Interprétation ionique
III.1 Le potentiel de repos
III.1.1 Données expérimentales
Des études menées sur des fibres nerveuses « géantes » de calmar, donnent la répartition ionique suivante
$$\begin{array}{|c|l|c|r|} \hline \text{Ions}& &\text{Concentration en }10^{3}\,mol/l&\\ \hline &\text{cytoplasme}&\text{Milieu extracellulaire}&\text{Eau de mer}\\ \hline K^{+}&400&20&10\\ \hline Na^{+}&50&440&460\\ \hline \end{array}$$
Ces résultats montrent une répartition ionique très inégale de part et d'autre de la membrane de l'axone : $Na$ et $(Cl)$ sont très peu abondants dans le milieu intracellulaire ; $K$ en revanche, y est vingt fois plus concentré que dans le milieu extracellulaire.
Le potentiel de membrane est lié à ces différences importantes de concentration de particules électriquement chargées.
III.1.2 Rôle des protéines membranaires
En raison de la différence de concentration entre le milieu intracellulaire et le milieu extracellulaire, les ions $K^{+}$ ont tendance à diffuser de l'intérieur vers l'extérieur de la cellule, alors que les ions $Na^{+}$ ont tendance à pénétrer dans la cellule.
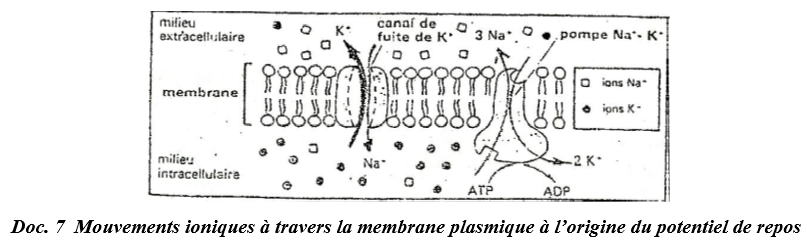
En réalité deux catégories de protéines membranaires prennent en charge le passage des ions à travers la bicouche lipidique : ce sont des « canaux » et des « pompes » à ions.
$\bullet\ $ Le « canal de fuite de $K^{+}$, ouvert en permanence, permet une diffusion passive des ions $K^{+}$ et de $Na^{+}$ dans le sens décroissant de leur gradient de concentration.
Ce canal étant beaucoup plus perméable aux ions $K^{+}$ qu'aux ions $Na^{+}$, il laisse donc « fuir » vers l'extérieur de la fibre beaucoup plus $K^{+}$ qu'il ne laisse entrer de $Na^{+}$, ce qui rend l'intérieur de la fibre électronégatif par rapport à l'extérieur.
$\bullet\ $ La « pompe $Na^{+}/K^{+}$ » assure le maintien de la dissymétrie ionique entre le milieu intracellulaire et le milieu extracellulaire.
Cette pompe est une enzyme protéique ($ATP$ ase) capable à la fois d'hydrolyser l'ATP et d'utiliser et d'utiliser l'énergie libérée pour assurer le transfert des ions $Na^{+}$ et $K^{+}$ contre leur gradient de concentration.
Une pompe $Na^{+}/K^{+}$ expulse plus de sodium dans le milieu extracellulaire qu'elle ne fait entrer de $K^{+}$ dans la fibre.
III.2 Le potentiel d'action
Des techniques expérimentales particulières ont permis de mettre en évidence au niveau de la membrane cytoplasmique, l'existence de canaux à $Na^{+}$ et à $K^{+}$ dont l'ouverture et la fermeture dépendent de la $ddp$ existante entre les deux faces de la membrane : ce sont les canaux voltage-dépendants.
A la suite de la stimulation d'une fibre nerveuse, on a suivi dans le temps, les variations de perméabilité membranaire aux ions $Na^{+}$ et $K^{+}$ (voir courbes ci-dessous).
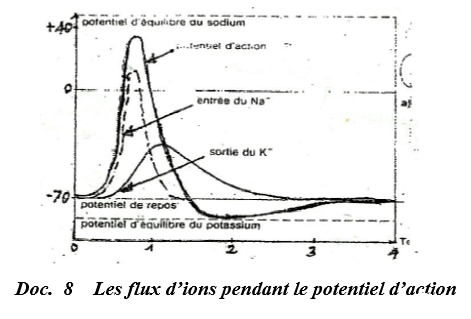
L'examen de la courbe révèle que la perméabilité de la membrane aux ions $Na^{+}$ augmente progressivement et atteint un maximum.
Au même moment, la dépolarisation de la fibre s'amplifie proportionnellement.
La repolarisation de la fibre s'accompagne d'une baisse de la perméabilité aux ions $Na^{+}$ et d'une augmentation progressive de cette perméabilité aux ions $K^{+}.$
Lorsque la sortie de $K^{+}$ atteint un maximum, la repolarisation est terminée.
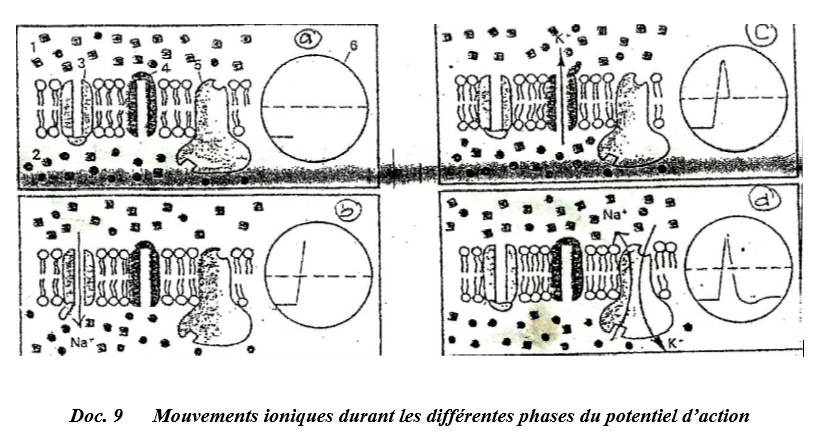
Il y a d'abord une augmentation rapide et importante de la perméabilité aux ions $Na^{+}$ qui s'engouffrent dans l'axone par ouverture des canaux $Na^{+}$ voltage-dépendants.
Cela entraine une suppression de charges positives dans l'axone et la face externe de la membrane devient électronégative par rapport à l'intérieur : c'est la dépolarisation (Doc. 19 b).
Puis, les canaux à $Na^{+}$ se ferment, inactivés, et les canaux à $K^{+}$ voltage-dépendants s'ouvrent.
Une sortie importante de $K^{+}$ permet la repolarisation (Doc. 19 c).
L'hyperpolarisation passagère est due à une sortie un peu plus importante de $K^{+}.$
Quand les canaux à $K^{+}$ se ferment, la polarisation est devenue normale, mais la répartition des ions reste anormale : excès de $Na^{+}$ interne et déficit de $K^{+}$ interne.
La pompe $Na^{+}/K^{+}$ augmente son activité pour rétablir la répartition normale en quelques millisecondes.
Tout blocage de la synthèse de l'ATP entraine l'arrêt de la pompe $Na^{+}/K^{+}$ (Doc. 19 d).
Remarques
$\bullet\ $ Le seuil de dépolarisation nécessaire pour déclencher un potentiel d'action correspond à la valeur de la dépolarisation pour ouvrir les canaux $Na^{+}$ voltage-dépendants.
$\bullet\ $ La période réfractaire qui succède au potentiel d'action et pendant laquelle la membrane est inexcitable, est due à l'inactivation de ces mêmes canaux à $Na^{+}$ ; cette inactivation correspond au temps nécessaire (quelques millisecondes) à la pompe $Na^{+}/K^{+}$ pour rétablir la répartition initiale.
$\bullet\ $ La « loi du tout ou rien » (constance de l'amplitude de $PA$) traduit le fait que cet évènement ne dépend que des concentrations ioniques de part et d'autre de la membrane d'une part, du nombre et des propriétés des différents canaux voltage-dépendants d'autre part.
Or, ces paramètres sont constants.
IV. Comparaison de potentiel d'action d'une fibre nerveuse et d'un nerf
On stimule une fibre isolée et un nerf avec des stimulations d'intensité croissante.
Le document suivant montre les résultats obtenus.
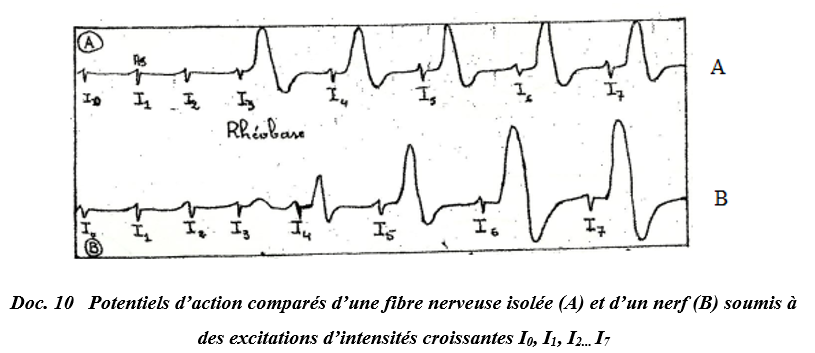
$-\ $ Au niveau de la fibre nerveuse, on obtient, dès que le seuil est atteint, un potentiel d'action d'amplitude d'emblée maximale et constante (quel que soit l'augmentation de l'intensité) :
On dit que la fibre obéit à la loi tout ou rien.
$-\ $ Dans le cas d'un nerf, l'amplitude augmente avec l'intensité des excitations puis demeure constante à partir d'une certaine intensité.
Une faible intensité touche un petit nombre de fibres.
Cette intensité augmentant, un nombre de fibres de plus en plus élevées est excité et l'amplitude devient constante lorsque toutes les fibres sont touchées.
On Parle ici d'un phénomène de recrutement.
Conclusion
Toute cellule nerveuse présente une polarité électrique due à une répartition inégale ionique.
L'inversion de cette polarité est à l'origine du potentiel d'action qui se propage.
Auteur:
Daouda Tine
Commentaires
Djibo Issa (non vérifié)
mar, 07/21/2020 - 11:50
Permalien
SVT
Fatima (non vérifié)
dim, 01/03/2021 - 20:34
Permalien
Terminal S2
Anonyme (non vérifié)
lun, 11/20/2023 - 04:59
Permalien
Ça devrait avoir un fichier
Ajouter un commentaire