
La reproduction chez les spermaphytes - Ts
Classe:
Terminale
Introduction
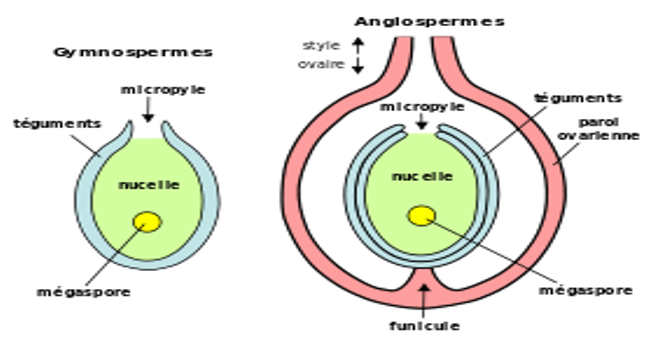
Beaucoup de végétaux possèdent des fleurs et des graines ; on leur a donné le nom de Phanérogames, « mariage visible », pour indiquer que la reproduction s'effectue dans un organe apparent, la fleur. La graine étant l'organe caractéristique, on préfère aujourd'hui le terme de spermaphytes, « plantes à graines ». Il faut alors distinguer deux groupes : les Gymnospermes, « plantes à graines nues », c'est-à-dire non enfermées dans les fruits, et les Angiospermes, « plantes à graines enveloppées », plus précisément enfermées dans des fruits.
On distingue les monocotylédones et les dicotylédones. Fleurs, fruits, graines sont les étapes essentielles de la reproduction.
I. Morphologie d'une fleur d'angiosperme
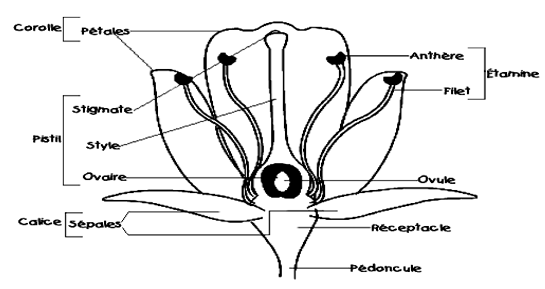
La fleur est constituée de deux pièces : des pièces stériles et des pièces fertiles.
Les pièces stériles forment le périanthe (enveloppe florale). Elles comportent :
$-\ $Les pétales qui donnent leur couleur à la fleur dont l'ensemble forme la corolle. Leur nombre est de $3$ ou multiple de $3$ chez les monocotylédones ; mais $5$ ou multiple de $5$ chez les dicotylédones.
$-\ $Les sépales sont de couleur verte et constituent les pièces les plus externes, l'ensemble forme le calice.
Les pièces fertiles sont réparties en androcée (partie mâle) et gynécée (partie femelle) :
$-\ $L'androcée forme l'ensemble des étamines dont chacune comprend un filet et une anthère.
$-\ $Le gynécée ou pistil est formé par les carpelles dont chacune présente à la base un ovaire renflé surmonté d'un style qui porte à son sommet un stigmate.
Remarque :
la plupart des angiospermes ont des fleurs hermaphrodites, de telles plantes sont dites hermaphrodites (bisexuées). Mais les fleurs peuvent être mâle ou femelle. Si une même plante porte à la fois des fleurs mâles et femelles, elle est dite dioïque. Si elle ne porte à la fois des fleurs mâles ou femelles, elle est dite monoïque.

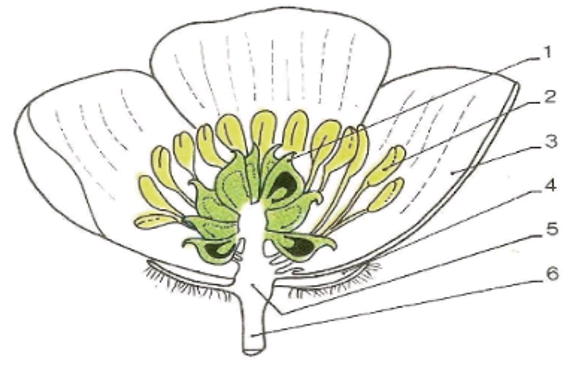
$1=n$ carpelles $\rightarrow$ gynécée ou pistil.
$2=n$ étamines $\rightarrow$ androcée.
$3=5$ pétales $\rightarrow$ corolle.
$4=5$ sépales $\rightarrow$ calice.
$5=$ réceptacle.
$6=$ pédoncule.
Document 1 : coupe longitudinale de la fleur
II. Étude des organes reproducteurs et gamètes
II.1. Étude de l'étamine
Chaque étamine comprend une partie mince, le filet, surmontée de l'anthère. Une coupe transversale dans une jeune anthère montre $4$ sacs polliniques. L'anthère s'ouvre par une fente longitudinale (fente de déhiscence). Les bords de la fente s'enroulent vers l'extérieur, exposant ainsi les grains de pollen.
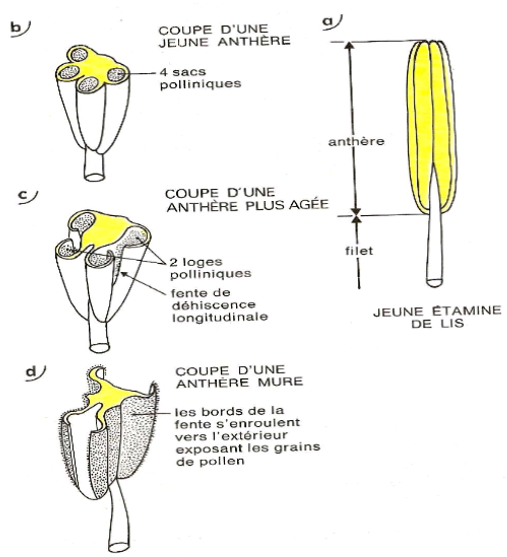
Document 2 : la structure de l'étamine
II.1.1. Anthère
L'anthère est formée de $2$ loges polliniques séparées par un sillon profond appelé connectif. Chaque loge pollinique est formée de $2$ sacs polliniques séparés par un sillon appelé fente de déhiscence : c'est par là que s'ouvrira l'anthère pour libérer le gamète mâle appelé grain de pollen.
L'observation microscopique d'un sac pollinique montre des cellules d'aspects différents et disposées en couche concentrique. Il s'agit de l'extérieur vers l'intérieur de l'épiderme à stomates, de la future assise mécanique, les deux assises nourricières et la masse centrale des cellules mères du pollen. Dans la zone du connectif, on distingue un faisceau conducteur et un parenchyme
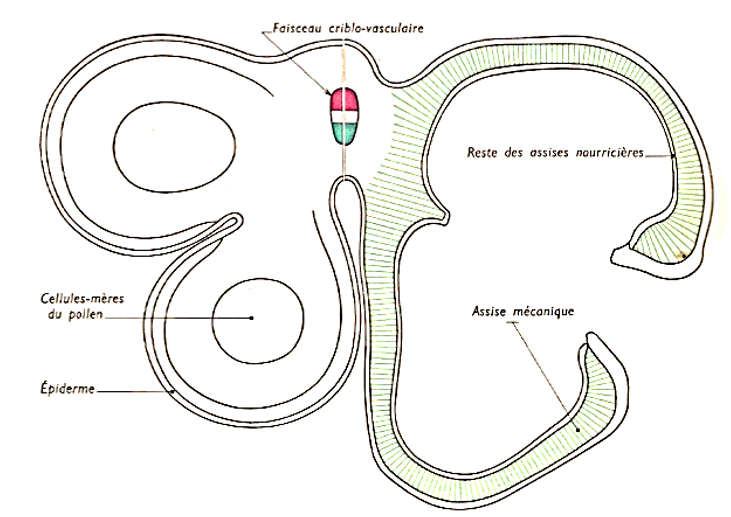
Coupe schématique d'une anthère de Lis
A gauche, $1/2$ coupe d'une anthère jeune ;
à droite, $1/2$ coupe d'une anthère mûre.
Document 3 : Coupe d'une Anthère
II.1.2. Le grain de pollen
II.1.2.1. Formation du pollen
La formation du grain de pollen comporte :
$-\ $une division réductionnelle qui permet d'obtenir deux cellules haploïdes réunies ensemble pour former une diade ;
$-\ $une division équationnelle qui permet d'obtenir $4$ petites cellules haploïdes ou microspores réunies pour former une tétrade.
$-\ $une division par mitose du noyau haploïde de chaque microspores pour former un grain de pollen.
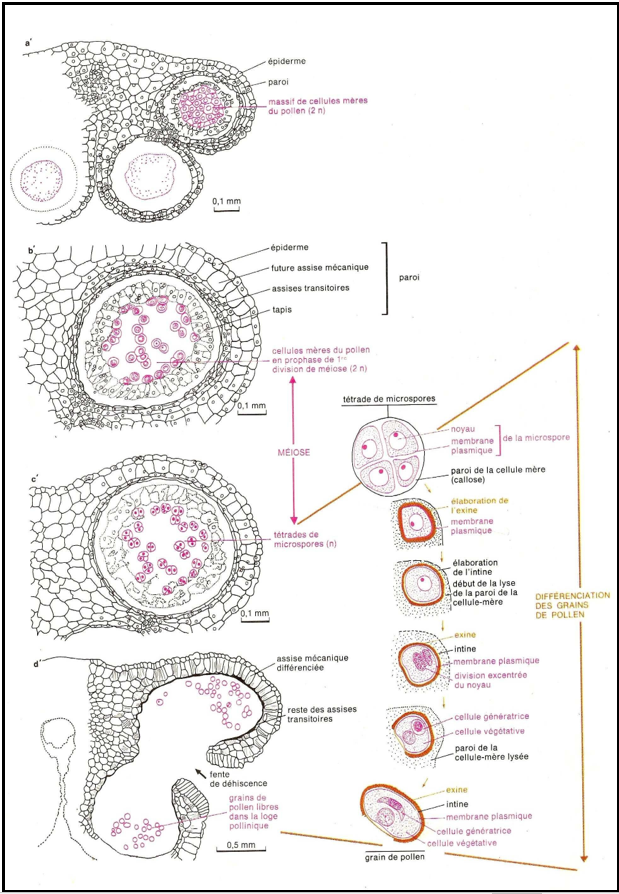
Document 4 : Formation du grain de pollen
II.1.2.2. Structure du grain de pollen
Le grain de pollen a une forme sphérique et il est constitué d'une paroi et d'un contenu cellulaire. La taille et l'ornementation des grains sont caractéristiques d'une espèce.
La paroi est formée de deux enveloppes : une enveloppe externe cutinisée et garnies d'épines appelée exine ; et une enveloppe interne cellulosique présentant des épaississement appelée intine.
Le contenu cellulaire est formé de deux cellules : une grande cellule dite végétative qui entoure une plus petite dite reproductrice, génératrice ou spermatogène
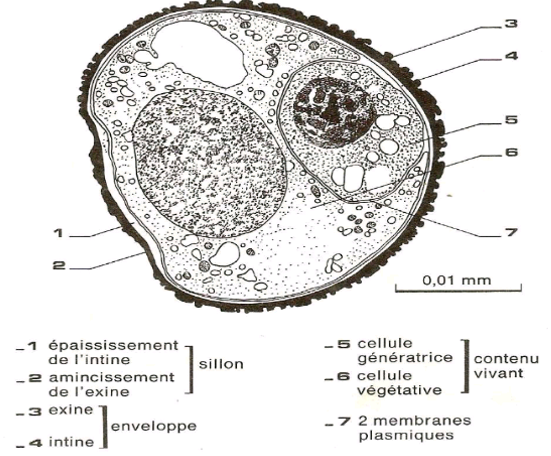
Document 5 : Coupe de grain de pollen
II.1.2.3. De la microspore au grain de pollen
Chaque microspore élabore une paroi externe complexe, l'exine, puis une deuxième paroi, l'intine, pectocellulosique, qui se place donc entre l'exine et la membrane plasmique.
Elle subit ensuite une division ; le noyau se déplace à la périphérie, se divise, puis le cytoplasme élabore une cloison incurvée qui isole une cellule de grande taille, la cellule végétative, et une cellule plus petite, la cellule génératrice (ou spermatogène). La cellule génératrice reste dépourvue de réserves contrairement à la cellule végétative qui accumule des réserves variées dans son cytoplasme (serviront en partie à la construction du tube pollinique). Ultérieurement, la cellule génératrice pénètre dans la cellule végétative (où elle est alors totalement incluse) puis prend une forme allongée.
A la fin de cette évolution, la paroi qui unissait la tétrade est hydrolysée ; les microspores devenues grains de pollen sont libérées dans la cavité du sac pollinique.
II.2. Étude du carpelle et ovule
II.2.1. Structure du carpelle (ovaire)
Le carpelle est formé de bas en haut d'une partie renflée, l'ovaire, prolongée par un axe, le style, terminé lui-même par une région papilleuse, le stigmate. Les carpelles sont soudés par les ovaires et par les styles ; seuls les stigmates sont distincts permettent de dénombrer les trois carpelles formant le pistil. Sa section triangulaire montre trois loges renfermant chacune deux rangées de masses plus ou moins sphériques, les ovules attachés par un pied, ou funicule, à l'axe de l'ovaire formant le placenta.
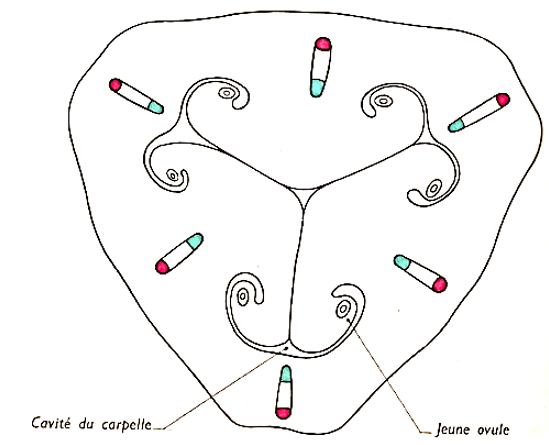
Document 6 : Coupe transversale schématique d'un jeune ovaire de Lis
II.2.2. L'ovule
II.2.2.1. Structure de l'ovule
L'observation microscopique montre les différentes parties suivantes :
$-\ $le funicule se prolonge sur le côté de l'ovule proprement dit et constitue le raphé. La limite finicule-raphé est appelée hile ;
$-\ $deux téguments étroitement accolés entourent l'ovule. A une extrémité, ils sont percés d’un orifice, le micropyle ;
$-\ $au centre, une masse cellulaire, le nucelle, abrite une grosse cellule claire accompagnée à chacune de ses extrémités de trois cellules ; l'ensemble forme le sac embryonnaire.
N.B :
On appelle chalaze la surface au niveau de laquelle nucelle et téguments se confondent (pôle opposé au micropyle). Il s'agit d'un ovule renversé.
Selon la forme de l'ovule par rapport au funicule, on distingue $3$ types d'ovules :
$-\ $l'ovule droit ou orthotrope dont le micropyle est dans l'axe du funicule ;
$-\ $l'ovule recourbé ou campylotrope dont le micropyle et le funicule forment un angle d'environ $90^{\circ}$ ;
$-\ $l'ovule inversé ou anatrope dont le micropyle et le funicule forment un angle de $180^{\circ}.$
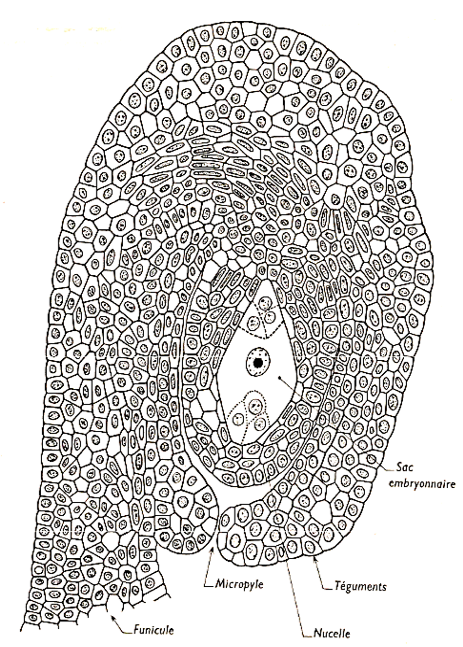
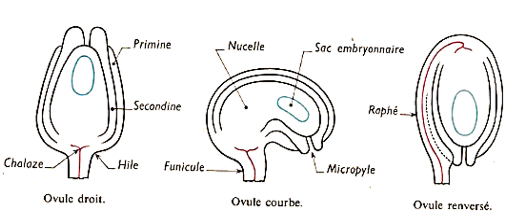
Document 7 : Coupe longitudinale d'un ovule $(A)$ ; trois types d'ovules $(B)$
II.2.2.2. Formation de l'ovule et du sac embryonnaire
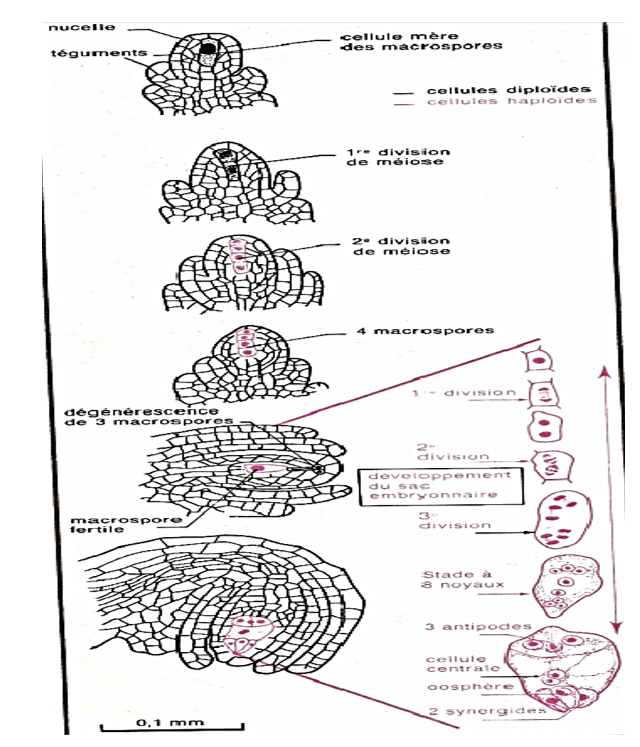
Le sac embryonnaire représente la partie essentielle de l'ovule. Dans l'ovule jeune, il n'est formé que d'une grosse cellule située dans le nucelle au voisinage du micropyle.
Cette cellule subit une première division réductionnelle suivie d'une division équationnelle ; il s'agit donc d'une méiose. On a alors quatre cellules à n chromosomes : les macrospores : trois dégénèrent et la quatrième grossit. Trois nouvelles divisions font apparaître $8$ noyaux répartis dans $7$ cellules de la manière suivante chez le Lis :
$-\ $Vers le micropyle, et dans l'axe du sac embryonnaire, l'oosphère flanquée de deux synergides
$-\ $Au pôle opposé, trois cellules, les antipodes ;
$-\ $Au centre, une cellule à deux noyaux secondaires ou noyaux du sac. Tous les noyaux sont à n chromosomes, en particulier l'oosphère ou gamète femelle.
La microscopie électronique permet de préciser les relations qui existent entre les cellules, en particulier au pôle micropylaire :
$-\ $l'oosphère est entourée généralement par les synergides ;
$-\ $celles-ci présentent à leur partie supérieure des replis qui interviendraient lors de la fécondation ; la cellule centrale enveloppe en grande partie oosphère et synergides
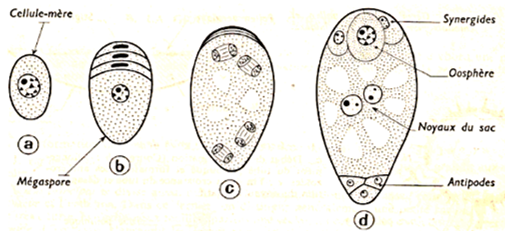
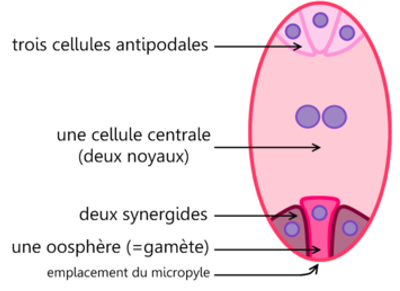
Document 8 : Mode de formation du sac embryonnaire
III. De la fleur au fruit
Elle comporte 3 étapes : la pollinisation, la germination et la croissance du tube pollinique, et la double fécondation.
III.1. La pollinisation
Elle correspond au transport de grains de pollen d'une anthère déhiscente jusqu'à leur dépôt sur un stigmate arrivé à maturité. Nous avons vu que la majorité des fleurs est bisexuée. Le pollen libéré par l'anthère peut donc être facilement transporté de l'étamine sur le stigmate de la même fleur (pollinisation directe).
Dans le cas où les fleurs sont unisexuées, le pollen d'une fleur est obligatoirement déposé sur le stigmate d'une autre fleur (pollinisation indirecte). Si les fleurs unisexuées sont portées par le même pied (espèce monoïque), on se retrouve dans les conditions peu différentes de la pollinisation directe. Si au contraire elles sont portées par des pieds différents (cas rare des espèces dioïques), une véritable pollinisation croisée est nécessaire. Le pollen étant dépourvu de mobilité propre, son transport est assuré par des agents extérieurs ; dans la plupart des cas ce sont le vent et les insectes qui assurent le transport :
$-\ $les plantes pollinisées par le vent sont appelées plantes anémophiles ;
$-\ $les autres espèces $(9/10$ environ$)$ sont pollinisées par les insectes (plantes entomophiles).
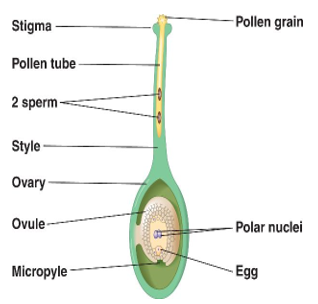
III.2 La germination du grain de pollen et la croissance du tube pollinique
III.2.1. La germination
L'eau contenue dans les sécrétions des cellules stigmatiques permet l'hydratation de grain de pollen. Ce qui entraîne l'émission de plusieurs d'excroissances de l'intine à travers les pores de l'exine. Parmi elles, une seule va s'allonger : c'est le tube pollinique
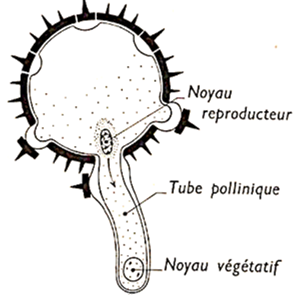
Document 9 : Tube pollinique
III.2.1. Orientation et croissance du tube pollinique
Les tubes polliniques des différents grains de pollen en s'allongeant s'engagent à travers des canaux fins situés dans le style, en direction des ovules situés dans l'ovaire généralement par le micropyle.
La croissance du tube est essentiellement une synthèse de paroi par le cytoplasme localisé à l'extrémité.
Cette croissance est possible :
$-\ $au début, grâce aux réserves fournies par la cellule végétative ;
$-\ $par la suite, grâce aux éléments fournis par les sécrétions ou les cellules gélifiées du style.
III.2.3. La formation des spermatozoïdes
Dès le début de la germination, le noyau de la cellule végétative s'engage dans le tube pollinique suivi de la cellule génératrice. Au cours de la traversée du style, la cellule génératrice se divise en deux spermatozoïdes ou anthérozoïdes
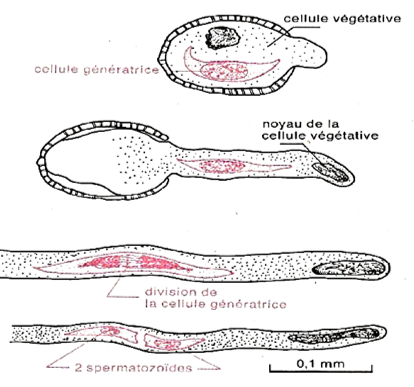
Document 10 : Formation des spermatozoïdes chez le Lis
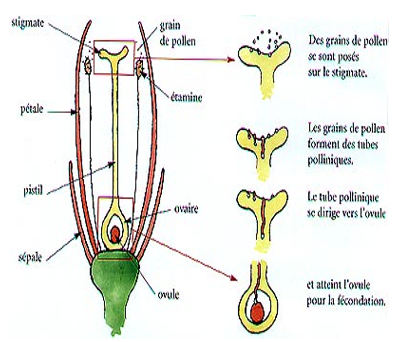
III.3. La fécondation
Arrivée contre le micropyle, le tube pollinique s'insinue entre les cellules de nucelle et arrive au contact du sac embryonnaire. Les observations récentes effectuées au microscope électronique ont précisé le déroulement des phases suivantes.
Le tube pollinique pénètre dans les replis d'une synergide et « décharge » ses $2$ spermatozoïdes à l'intérieur. Après la traversée de la synergide, les $2$ gamètes sont déversés :
$-\ $L'un au voisinage de l'oosphère ;
$-\ $l'autre au voisinage de la cellule centrale.
Le premier spermatozoïde « fusionne » avec l'oosphère, les noyaux s'unissent, un œuf diploïde est constitué (œuf principal).
Le deuxième spermatozoïde « fusionne » avec la cellule centrale, son noyau s'unit avec le noyau diploïde de celle-ci ; un deuxième œuf, triploïde, est formé, l'œuf accessoire. Il y a donc double fécondation.
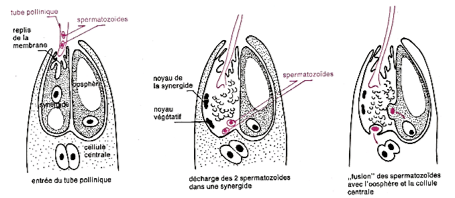
Document 11 : la double fécondation
IV. Formation de la graine et du fruit
A la suite de la fécondation, la fleur se transforme en fruit (c'est généralement l'ovaire qui constitue la partie principale du fruit), le (ou les) ovule(s) en graine(s).
IV.1. Formation de la graine
$-\ $Évolution de l'œuf principal et de l'œuf accessoire.
Après la fécondation, l'œuf principal se divise et engendre l'embryon. D'abord globuleux, l'embryon prend ensuite une forme en cœur dans laquelle on reconnaît :
$-\ $une ébauche de racine (la radicule)
$-\ $une ébauche de bourgeon terminal (la gemmule)
$-\ $une ébauche de (ou des) cotylédons (s).
La zone située entre la radicule et l'attache des cotylédons est la tigelle. Parallèlement, l'œuf accessoire se divise également et donne un massif cellulaire, l'albumen qui entoure l'embryon. Albumen et embryon se développent ensuite côte à côte mais à des vitesses variables suivant les espèces.
$\blacktriangleright\ $La transformation des téguments ovulaires.
Le (ou les) tégument (s) de l'ovule deviennent le (ou les) tégument (s) de la graine, tandis que le micropyle se ferme. Ils sont plus ou moins épais et diversement colorés.
$\blacktriangleright\ $L'accumulation des réserves.
Au cours du développement de l'embryon et de l'albumen, des réserves variées s'accumulent soit dans l'albumen (graines à albumen), soit dans les cotylédons (graines sans albumen).
Ainsi la graine mûre est un organe formé d'un embryon, de réserves, protégés par les téguments ; l'ensemble étant en vie ralentie. La graine se détache du funicule qui laisse une cicatrice, le hile

Document 12 : formation de la graine
IV.2. Formation du fruit
Après la maturation des graines issues de la transformation des ovules fécondées, les carpelles se transforment. Ces transformations concernent :
$-\ $le parenchyme qui peut se charger de substances alimentaires comme dans le cas des fruits charnus ;
$-\ $l'épiderme à stomates devient l'enveloppe du fruit ;
$-\ $le stigmate, le style et le reste de l'androcée dégénèrent.
Remarque :
Chez les angiospermes, les graines sont complètement contenues dans le fruit
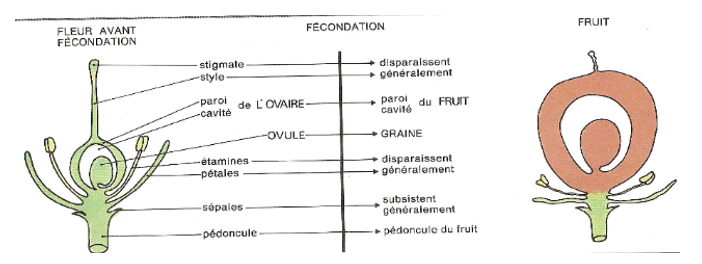
Document 13 : De la fleur au fruit
V. Cycle de développement et chromosomique
La fleur portant les organes reproducteurs donne naissance aux gamètes mis en contact par la pollinisation et la germination des grains de pollen. Ces œufs issus de cette double fécondation se développent pour donner la graine. Après la germination cette graine donne un appareil végétatif donnant lieu à une plante qui sera à l'origine de nouvelles fleurs : c'est le cycle de développement.
Dans le cycle de développement d'un spermaphyte, on retrouve la succession de deux phases :
$-\ $une phase diploïde à $2n$ chromosome ou diplophase représentée par l'œuf, la graine, la plante feuillée. Cette phase se termine par la production des spores (microspores et macrospores) : la plante feuillée, fleurie est donc un sporophyte.
$-\ $Une phase haploïde à n chromosomes ou haplophase représentée par les grains de pollen et le sac embryonnaire. Ces organes produisent des gamètes : ce sont des gamétophytes. Chez tous les êtres vivants se succèdent ainsi deux phases. Les spermaphytes ont donc un cycle haplo-diplophasique.
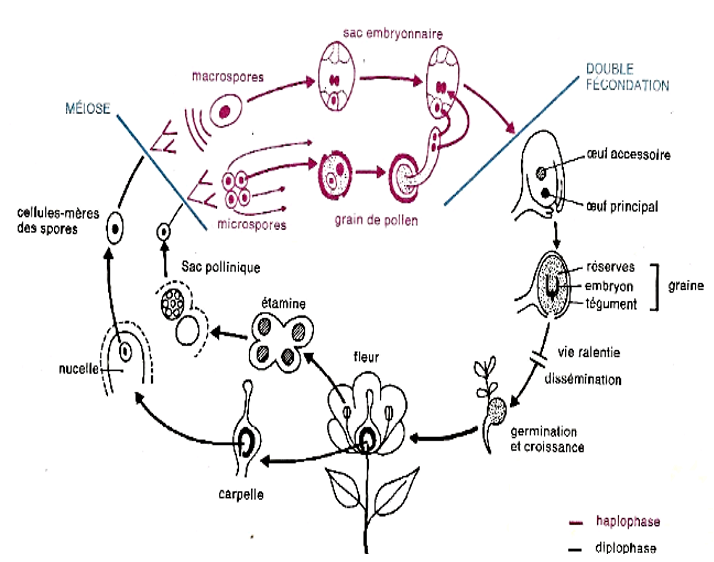
Document 14 : Cycle de développent d'une angiosperme à fleurs bisexuées
Auteur:
Daouda Tine
Commentaires
Anonyme (non vérifié)
sam, 01/07/2023 - 22:40
Permalien
Très bien
BIVIHOU IWANGOU... (non vérifié)
sam, 05/25/2024 - 21:12
Permalien
Appreciation
hubert (non vérifié)
mar, 06/03/2025 - 19:28
Permalien
j'aimerais bien téléchargé ce cours
el malick (non vérifié)
jeu, 07/31/2025 - 19:34
Permalien
svt
Anonyme (non vérifié)
lun, 08/25/2025 - 11:21
Permalien
Très bien
Ajouter un commentaire