
Corrigé Bac svt 1er groupe S2-S2A-S4-S5 2018
I. Maitrise des connaissances
Introduction
La moelle épinière est le centre nerveux auquel sont rattachés les nerfs rachidiens.
Le sujet soumis à notre réflexion pose le problème de son organisation, de celle d'un nerf rachidien et des rapports entre ces deux structures anatomiques. Par un exposé structuré et illustré, nous allons d'abord rappeler l'organisation au microscope de la moelle épinière et celle d'un nerf rachidien puis l'expérience de Waller qui montre la structure anatomique entre ces deux structures.
A. Structure de la moelle épinière et du nerf rachidien.
1. Structure de la moelle épinière
a. Préparation de substance grise observée au microscope optique.
$\centerdot\ \ $ La substance grise observée au microscope optique présent des péricaryons, des fibres nerveuses et des cellules gliales.
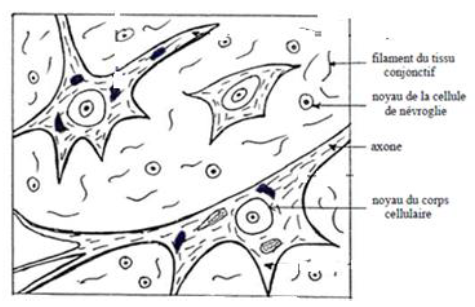
Préparation de substance grise observée au microscope.
b. Préparation de substance blanche observée au microscope optique.
$\centerdot\ \ $ La substance blanche médullaire est formée de fibres nerveuses entourées chacune d'une gaine de myéline interrompue au niveau des nœuds de ravier
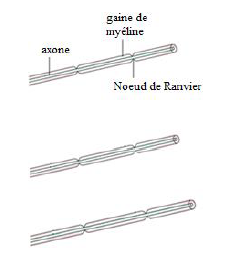
Structure de la substance blanche médullaire observée au microscope optique
2. Structure du nerf rachidien
Un nerf rachidien dilacéré puis observé au microscope présente des fibres nerveuses entourées chacune d'une gaine de myéline doublée d'une gaine de Schwann.
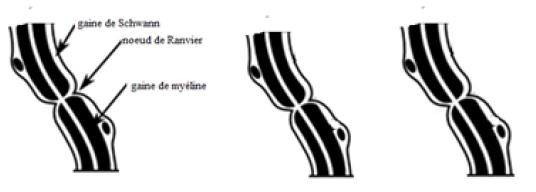
Structure du nerf rachidien
B. Expériences de dégénérescence Walllérienne
La section d'un nerf rachidien d'un animal entraine après quelques jours la dégénérescence des fibres dans le bout périphérique alors que les fibres du bout central survivent.
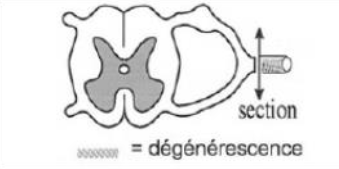
Cette expérience prouve qu'il y a une continuité anatomique entre corps cellulaire et fibre nerveuse. L'ensemble forme une seule unité structurale qui est la cellule nerveuse ou neurone.
II. Compétences méthodologiques
Exercice 1
1. Les documents 1 et 2 montrent que l'antigène Rhésus est gouverné par un gène autosomique ayant deux (2) allèles dont l'un est dominant.
$Rh^{+}\ $ est l'allèle dominant ; $Rh^{-}\ $ est l'allèle récessif.
$-\ $ Un sujet $[Rh^{-}]$ fabrique des anticorps contre les hématies $[Rh^{+}]$ lorsque ces dernières pénètrent dans son organisme.
$-\ $ Le premier enfant du couple a pour phénotype $[Rh^{-}].$ Il a le même phénotype que sa mère madame $X$ qui ne produit pas des anticorps $\text{anti-}Rh^{-}.$
Le phénotype de cet enfant montre que son père est hétérozygote pour le facteur Rhésus. Le génotype du père est donc $Rh^{+}//Rh^{-}.$
Le second enfant du couple a le phénotype $[Rh^{+}].$ Sa mère madame $X$ produit des anticorps $\text{anti-}Rh^{+}$ si les hématies de cet enfant pénètrent dans son organisme.
$-\ $ Les trois petites sœurs du second enfant sont mortes avant leurs naissances (mort-nées)
Le document 3 montre que la mère $[Rh^{-}]$ ayant un premier fœtus $[Rh^{+}]$ ne fabrique pas d'anticorps $\text{anti-}Rh^{+}$ (IgD) durant toute la grossesse à cause d'une absence de contact entre son sang et celui de l'enfant qu'elle porte. Cependant, à partir de l'accouchement, la mère produit une quantité croissante d'IgD. Cette production d'IgD s'explique par le contact entre le sang de la mère et celui de l'enfant lors de l'accouchement. Ces IgD permettent l'immunisation de l'organisme maternel contre les antigènes Rhésus.
Le document 4 montre que les IgD maternelles peuvent traverser le placenta et provoquer l'agglutination des hématies du fœtus de phénotype $[Rh^{+}].$ Les complexes-immuns ainsi formés activent la destruction des hématies. Cela aboutit à la mort du fœtus.
Bilan.
Les hématies portant l'antigène Rhésus du premier enfant $[Rh^{+}]$ de madame $X$ sont entrées en contact avec l'organisme maternel au cours de l'accouchement et ont provoqué la production d'IgD.
Les trois petites sœurs de cet enfant $[Rh^{+}]$ étant aussi $[Rh^{+}]$, les IgD maternelles ont traversé le placenta et ont provoqué à chaque fois l'agglutination puis la destruction de leurs hématies. Cette maladie hémolytique explique les trois morts successives avant les naissances.
2. Madame $X\;,\ [Rh^{-}]$, a le génotype $Rh^{-}//Rh^{-}.$
Monsieur $X\;,\ [Rh^{+}]$ ayant un fils $[Rh^{-}]$ est donc de génotype $Rh^{+}//Rh^{-}.$
$$\begin{array}{lcc}&\text{Madame }X&\text{Monsieur }X\\ \\ \text{Génotype des parents :}&Rh^{-}//Rh^{-}&Rh^{+}//Rh^{-}\\ \\ \text{Gamètes}&Rh^{-}/&Rh^{+}/\;;\ Rh^{-}/\end{array}$$
Échiquier de croisement
$$\begin{array}{|c|c|c|}\hline\text{Gamètes}&Rh^{+}/&Rh^{-}/ \\ \hline Rh^{-}/&Rh^{+}//Rh^{-}[Rh^{+}]&Rh^{-}//Rh^{-}[Rh^{-}]\\ \hline\end{array}$$
Ce couple peut donc avoir d'autres enfants $[Rh^{-}].$ Ces derniers n'ayant pas l'antigène Rhésus sur leurs hématies seront sains car les IgD maternelles n'agglutinent pas les hématies dépourvues d'antigène Rhésus.
Exercice 2
1. a) On étudie deux caractères :
$-\ $ La longueur du poil,
$-\ $ La couleur du poil.
On s'aperçoit que, pour les deux croisements, tous les individus $F_{1}$ ont le même phénotype pour la longueur des poils : ras. Donc le gène déterminant ce caractère est porté par les autosomes.
b) L'allèle responsable du phénotype "ras" est dominant.
c) On écrit : allèle déterminant le phénotype dominant : $R$ ; allèle déterminant le phénotype récessif : $I$
En revanche, pour l'autre caractère, en l'occurrence la couleur des poils, les résultats en $F_{1}$ sont différents selon le croisement étudié.
Les mâles ont toujours le phénotype de leur mère, les femelles ont un phénotype intermédiaire.
Le gène déterminant ce caractère est porté par les chromosomes sexuels. Cela ne peut être $Y$ car le caractère est présent chez les mâles et les femelles. C'est donc le gonosome $X$ qui porte le gène.
Les femelles qui ont deux chromosomes $X$ ont deux allèles du gène ; en $F_{1}$, elles ont hérité d'un allèle différent de chaque parent et on observe qu'elles sont bicolores. Les deux allèles s'expriment simultanément : ils sont codominants. On les notera $N\ $ et $\ O.$
2- Premier croisement
On peut écrire les phénotypes et les génotypes des individus du 1er croisement :
$$\begin{array}{lccc}\text{Phénotypes :}&\text{mâle }[R\;,\ N]&\times&\text{femelle }[I\;,\ O]\\ \\ \text{Génotypes :}&R//RX^{N}Y&\times&I//IX^{O}X^{O}\end{array}$$
Le mâle produit deux types de gamètes : $R, X^{N}\;;\ R, Y.$ La femelle ne produit qu'un type de gamètes : $I, X^{O}.$
$$\begin{array}{|c|c|c|}\hline\gamma\ \text{ mâle}\backslash\gamma\ \text{ femelle}&R, X^{N}\;50\%&R, Y\;50\%\\ \hline I, X^{O}\;100\%&R//IX^{N}X^{O}\;50\%&R//IX^{O}Y\;50\%\\ \hline\end{array}$$
Cela est conforme aux résultats : les mâles sont orangés, à poils ras, les femelles sont bicolores à poils ras.
$-\ $ Deuxième croisement
De même, on peut écrire pour le 2e croisement :
$$\begin{array}{lccc}\text{Phénotypes :}&\text{mâle }[I\;,\ O]&\times&\text{femelle }[R\;,\ N]\\ \\ \text{Génotypes :}&I//IX^{O}Y&\times&R//RX^{N}X^{N}\end{array}$$
Échiquier de croisement
$$\begin{array}{|c|c|c|}\hline\gamma\ \text{ mâle}\backslash\gamma\ \text{ femelle}&I, X^{O}\;50\%&I, Y\;50\%\\ \hline R, X^{N}&R//IX^{O}X^{N}&R//IX^{N}Y\\ \hline\end{array}$$
En $F_{1}$, les cobayes mâles sont bien noirs à poils ras, et les cobayes femelles bicolores à poil ras.
2. Si l'éleveur croise un mâle de génotype $R//IX^{O}Y$ avec une femelle de génotype $R//IX^{O}X^{N}$ il obtiendra une descendance dont les résultats sont consignés dans le tableau ci-dessous :
$$\begin{array}{|c|c|c|c|c|}\hline\gamma\text{ mâle}\backslash\gamma\text{ femelle}&R/, X^{O}&I/, X^{O}&R/, Y&I/, Y\\ \hline R/, X^{N}&R//RX^{O}X^{N}&R//IX^{O}X^{N}&R//RX^{N}Y&R//IX^{N}Y\\ \hline R/, X^{O}&R//RX^{O}X^{O}&R//IX^{O}X^{O}&R//RX^{O}Y&R//IX^{O}Y\\ \hline I/, X^{N}&R//IX^{O}X^{N}&I//IX^{O}X^{N}&R//IX^{N}Y&I//IX^{N}Y\\ \hline I/, X^{O}&R//IX^{O}X^{O}&I//IX^{O}X^{O}&R//IX^{O}Y&I//IX^{O}Y\\ \hline\end{array}$$
Si l'éleveur croise un mâle de génotype $R//IX^{N}Y$ avec une femelle de génotype $R//IX^{O}X^{N}$ il obtiendra la descendance dont le génotype figure dans le tableau suivant.
$$\begin{array}{|c|c|c|c|c|}\hline\gamma\text{ mâle}\backslash\gamma\text{ femelle}&R/, X^{N}&I/, X^{N}&R/, Y&I/, Y\\ \hline R/, X^{N}&R//RX^{N}X^{N}&R//IX^{N}X^{N}&R//RX^{N}Y&R//IX^{N}Y\\ \hline R/, X^{O}&R//RX^{O}X^{N}&R//IX^{O}X^{N}&R//RX^{O}Y&R//IX^{O}Y\\ \hline I/, X^{N}&R//IX^{N}X^{N}&I//IX^{N}X^{N}&R//IX^{N}Y&I//IX^{N}Y\\ \hline I/, X^{O}&R//IX^{O}X^{N}&I//IX^{O}X^{N}&R//IX^{O}Y&I//IX^{O}Y\\ \hline\end{array}$$
3. L'éleveur ne pourra pas obtenir de cobayes mâles bicolores à poils longs puisque les mâles n'ayant qu'un $X$, n'ont qu'un allèle du gène déterminant la couleur.
En revanche, dans les deux cas, il obtiendra dans la population de femelles 1/8, soit $12.5\%$ de cobayes femelles bicolores à poils longs qui possèdent un génotype $I//IX^{O}X^{N}$
Commentaires
Anonyme (non vérifié)
mer, 01/06/2021 - 20:44
Permalien
J'aime bien l'épreuve
Ajouter un commentaire