
Corrigé Bac SVT 1er Groupe S2 S2A S4 S5 2011
I. Maitrise des connaissances
Introduction
Dans l'organisme, les muscles squelettiques sont sous la commande des nerfs moteurs.
Le message nerveux qui parcourt le motoneurone traverse la plaque motrice puis déclenche la contraction musculaire.
Une synapse neuromusculaire est une jonction entre l'arborisation terminale d'un neurone moteur et une cellule musculaire.
Le franchissement de la synapse neuromusculaire par l'influx nerveux nécessite la libération d'un neurotransmetteur excitateur : l'acétylcholine.
La molécule de curare qui a la même conformation spatiale que l'acétylcholine occupe ses récepteurs sur l'appareil sous-neural.
Comment fonctionne la plaque motrice ?
comment un poison comme le curare peut-il perturber la transmission synaptique de l'influx nerveux à travers la synapse neuromusculaire ?
c'est à ces questions que nous tenterons de répondre dans notre exposé.
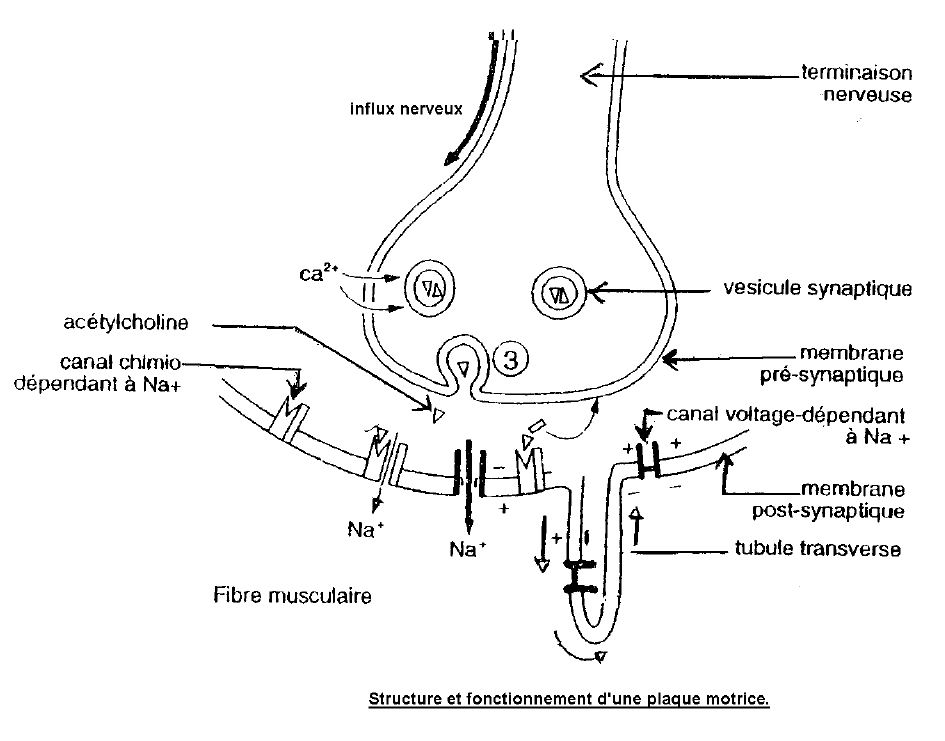
1) Le fonctionnement de la plaque motrice
L'arrivée du potentiel d'action au niveau de la membrane pré-synaptique déclenche la succession des événements suivants :
$-\ $ Entée d'ions $Ca^{++}$ dans la terminaison nerveuse qui entraine la libération d'acétylcholine dans la fente synaptique.
$-\ $ L'acétylcholine libéré se fixe sur les récepteurs de la membrane post-synaptique qui sont des canaux à $Na^{+}$ chimio-dépendants.
$-\ $ Les canaux à $Na^{+}$ chimio-dépendants s'ouvrent d'où une entée d'ions $Na^{+}$ dans la fibre musculaire dont la membrane se dépolarise.
$-\ $ Une enzyme l'acétylcholinestérase hydrolyse l'acétylcholine.
$-\ $ Choline issue de cette inactivation de l'acétylcholine est réabsorbée au niveau de la membrane pré-synaptique.
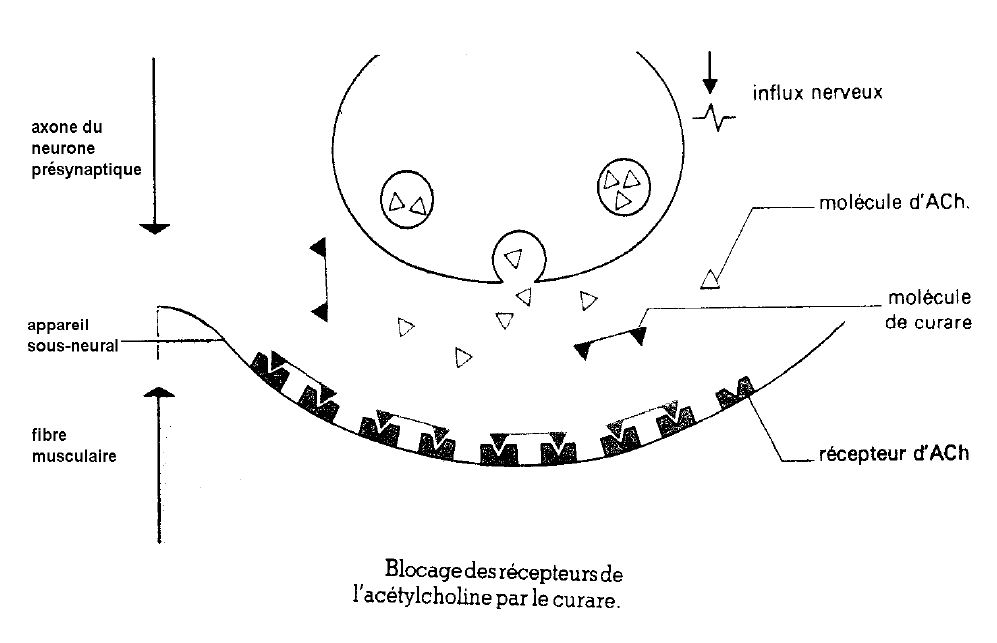
2) Perturbation par le curare de la transmission synaptique de l'influx nerveux
La molécule de curare, poison d'origine végétale, mime grossièrement à ses deux extrémités, une molécule d'acétylcholine.
Elle se fixe sur les récepteurs à acétylcholine de l'appareil sous-neural et provoque la paralysie.
Conclusion
L'arrivée du potentiel d'action au niveau du bouton synaptique déclenche la libération de l'acétylcholine dans la fente synaptique.
L'acétylcholine se fixe sur les récepteurs de la membrane post synaptique d'où une entrée de Na+ provoquant la dépolarisation de la fibre musculaire.
Le curare occupe les récepteurs à acétylcholine de la membrane musculaire, bloquant ainsi la transmission synaptique de l'influx nerveux.
II. Exploitation de documents
A)
1) Avant l'injection de sel de calcium ou d'un agent chélateur, la calcémie est normale.
L'injection de sel de calcium élève la calcémie à $130\%$ par rapport à la normale alors que l'injection d'un agent chélateur abaisse la calcémie à $85\%$ par rapport à la normale.
$2$ heures après les injections, la calcémie redevient normale et constante.
2) Puisque la calcémie redevient normale et constante au bout de $2$ heures après les perturbations, on en conclut que la calcémie est régulée.
B)
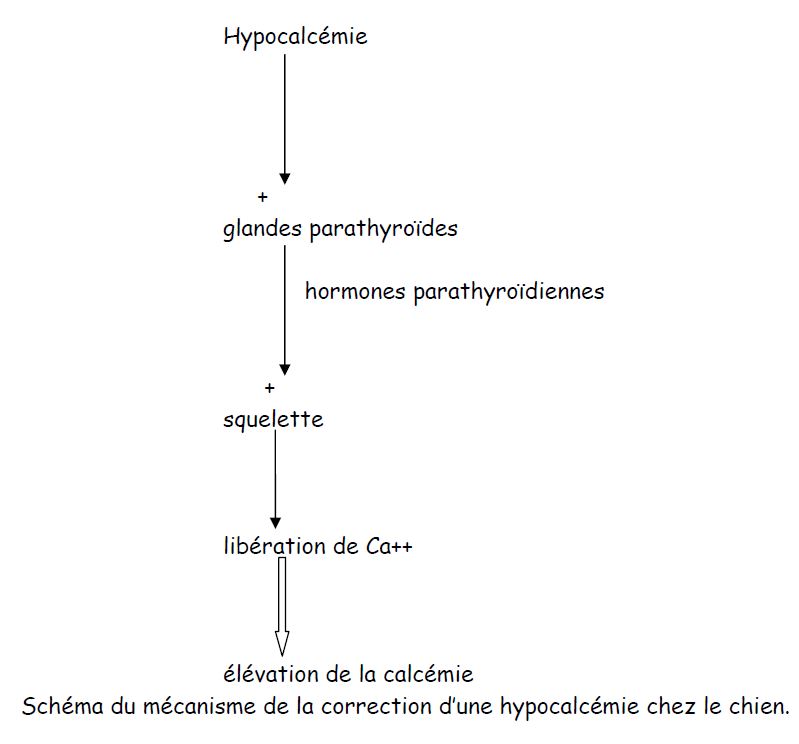
3) Dans les $2$ heures qui précédent l'ablation des glandes parathyroïdes, la calcémie est constante et égale à $100\;mg/l.$
Après ablation des glandes parathyroïdes, la calcémie diminue et passe de $95$ à $70\;mg/l$ de la $3e$ à la $11e$ heure.
4) Les glandes parathyroïdes semblent élever la calcémie.
5) L'injection d'extraits parathyroïdiens à un chien normal élève la calcémie qui passe de $100\;mg/l$ à $180\;mg/l$ au $4e$ jour.
Du $4e$ au $6e$ jour la calcémie revient à la valeur normale.
Du $6e$ au $8e$ jour elle passe au dessous de sa valeur normale.
Au-delà du $8e$ jour elle est constante et voisine de $100\;mg/l.$
Les glandes parathyroïdes élèvent la calcémie par l'intermédiaire d'une substance chimique véhiculée par le sang.
6) Les glandes parathyroïdes provoquent la libération de calcium par les os.
Le calcium libéré par les os élève la calcémie du milieu intérieur.
7) L'injection de sang ayant un faible taux de calcium à un chien normal élève la calcémie de ce dernier.
Donc le sang dont la teneur en calcium est inférieure à la normale déclenche, au contact de la glande parathyroïde, la sécrétion d'hormones parathyroïdiennes hypercalcémiantes.
8)
III. Pratique du raisonnement scientifique
Partie I :
Le croisement des canaris argentés donne $\dfrac{2}{3}$ de canaris argentés et $\dfrac{1}{3}$ de canaris dorés.
On en déduit que le caractère doré qui apparaît dans la descendance est récessif et les canaris argentés sont hybrides.
Les résultats statistiques de la descendance du croisement entre les canaris argentés s'expliquent par le fait que l'allèle responsable du caractère « argenté » est létal à l'état homozygote.
On pose :
A allèle responsable du caractère « argenté ».
d'allèle responsable du caractère « dorée ».
$1^{er}$ croisement :
Phénotype $[A]\times[A]$
Génotypes $\dfrac{\underline{A}}{d}\times\dfrac{\underline{A}}{d}$
gamètes $50\%\;\underline{A}\;;\ 50\%\;\underline{d}$
résultats
$$\begin{array}{|c|c|c|}\hline\Theta\backslash\Upsilon&\underline{A}&\underline{d}\\ \hline\underline{A}&\dfrac{\underline{A}}{A}\ \text{ Non viable}&\dfrac{\underline{A}}{d}[A]\\ \hline \underline{d}&\dfrac{\underline{A}}{d}[A]&\dfrac{\underline{d}}{d}[d]\\ \hline \end{array}$$
décompte des phénotypes
$\dfrac{2}{3}\;[A]$
$\dfrac{1}{3}\;[d]$
$2^{ème}$ croisement
Phénotype $[A]\times[d]$
Génotypes $\dfrac{\underline{A}}{d}\times\dfrac{\underline{d}}{d}$
gamètes $50\%\;\underline{A}\;;\ 50\%\;\underline{d}\qquad 100\%\;\underline{d}$
descendance
$$\begin{array}{|c|c|c|} \hline\Theta\backslash\Upsilon&A&d\\ \hline d&\dfrac{\underline{A}}{d}[A]&\dfrac{\underline{d}}{d}[d]\\ \hline \underline{d}&\dfrac{\underline{A}}{d}[A]&\dfrac{\underline{d}}{d}[d]\\ \hline \end{array}$$
décompte des phénotypes
$\dfrac{1}{2}\;[A]$
$\dfrac{1}{2}\;[d]$
Partie II
1) Phénotype agate $[n^{+}\ b]$
Phénotype isabelle $[n\ b]$
2) Dans le premier croisement la $F_{1}$ est homogène alors que dans le croisement réciproque, la $F_{1}$ est hétérogène (le phénotype des mâles est différent de celui des femelles).
Hypothèse :
Les gènes responsables du phénotype agate et du phénotype isabelle sont portés par un gonosome.
3) Lorsque le parent mâle est de phénotype agate, toute la descendance est homogène et de phénotype agate.
Lorsque le parent femelle est agate (phénotype dominant) la descendance est hétérogène.
Donc chez le canari, le mâle est homogamétique $ZZ$ et la femelle est hétérogamétique $ZW.$
$1^{er}$ croisement :
phénotypes des parents $\Upsilon\;[n^{+}\ b]\times\Theta\;[n\ b]$
Génotype des parents $\Upsilon\;\dfrac{\underline{Z_{b}^{n^{+}}}}{Z_{b}^{n^{+}}}\times\Theta\;\dfrac{\underline{Z_{b}^{n}}}{W}$
Gamètes des parents $100\%\;Z_{b}^{n^{+}}\qquad\dfrac{1}{2}Z_{b}^{n}\;;\ \dfrac{1}{2}W$
$F_{1}$ par l'échiquier
$$\begin{array}{|c|c|c|} \hline\Upsilon\backslash\Theta&Z_{b}^{n}&W\\ \hline Z_{b}^{n^{+}}&\dfrac{\underline{Z_{b}^{n^{+}}}}{Z_{b}^{n}}\;\Upsilon\;[n^{+}\ b]&\dfrac{\underline{Z_{b}^{n}}}{W}\;\Theta\;[n^{+}\ b]\\ \hline \end{array}$$
$2^{ème}$ croisement :
Phénotypes des parents : $\Theta\;[n^{+}\ b]\times\Upsilon\;[n\ b]$
Génotypes des parents : $\dfrac{\underline{Z_{b}^{n^{+}}}}{W}\times\dfrac{\underline{Z_{b}^{n}}}{Z_{b}^{n}}$
Gamètes des parents
$\dfrac{1}{2}\;Z_{b}^{n+}\;;\ \dfrac{1}{2}\;W\qquad 100\%\;Z_{b}^{n}$
$F_{1}$
$$\begin{array}{|c|c|c|} \hline\Upsilon\backslash\Theta&Z_{b}^{n^{+}}&W\\ \hline Z_{b}^{n}&\dfrac{\underline{Z_{b}^{n^{+}}}}{Z_{b}^{n}}\;\Upsilon\;[n^{+}\ b]&\dfrac{\underline{Z_{b}^{n}}}{W}\;\Theta\;[n^{+}\ b]\\ \hline \end{array}$$
B)
$-\ $ phénotypes $\Upsilon\;[n\ b]\times\Theta\;[n^{+}\ b^{+}]$
$-\ $ génotypes des parents $\Upsilon\;\dfrac{\underline{Z_{b}^{n}}}{Z_{b}^{n}}\times\Theta\;\dfrac{\underline{Z_{b^{+}}^{n^{+}}}}{W}$
gamètes des parents $100\%\;Z_{b}^{n}\qquad\dfrac{1}{2}\;Z_{b^{+}}^{n^{+}}\;;\ \dfrac{1}{2}\;W$
$$\begin{array}{|c|c|c|} \hline\Upsilon\backslash\Theta&Z_{b^{+}}^{n^{+}}&W\\ \hline Z_{b}^{n}&\dfrac{\underline{Z_{b^{+}}^{n^{+}}}}{Z_{b}^{n}}\;\Upsilon\;[n^{+}\ b^{+}]&\dfrac{\underline{Z_{b}^{n}}}{W}\;\Theta\;[n\ b]\\ \hline \end{array}$$
Décompte des phénotypes
$\dfrac{1}{2}\Upsilon\;[n^{+}\ b^{+}]$
$\dfrac{1}{2}\Theta\;[n\ b]$
C) $\Theta[n\ b\ d]\times\Upsilon\;[n^{+}\ b^{+}\ A]$
génotype de la $\Theta\ :\ \dfrac{\underline{Z_{b}^{n}}}{W}\;\dfrac{\underline{d}}{d}$
génotypes possibles du $\Upsilon\ :\ \dfrac{\underline{Z_{b^{+}}^{n^{+}}}}{Z_{b^{+}}^{n^{+}}}\;\dfrac{\underline{A}}{d}\quad\text{ou}\quad\dfrac{\underline{Z_{b^{+}}^{n^{+}}}}{Z_{b}^{n}}\;\dfrac{\underline{A}}{d}$
Commentaires
Ramatoulaye Ndiaye (non vérifié)
ven, 07/26/2024 - 00:18
Permalien
Bac
Ajouter un commentaire