
Rôle du système nerveux dans le comportement moteur d'un animal - Ts2
Classe:
Terminale
Introduction
Dans leur environnement les animaux sont soumis à de nombreux stimuli externes ou internes auxquels ils répondent par une succession de réactions appelées comportements. Le comportement moteur d'un animal correspond à deux types de mouvements :
$-\ \ $ Les mouvements involontaires et inconscients qu'on appelle mouvements réflexes ;
$-\ \ $ Les mouvements volontaires spontanés.
I. L'activité réflexe
I.1. Notion de réflexe
Le comportement moteur des animaux peut être déclenché sans l'intervention de la volonté, c'est l'activité réflexe, qui peut être innée (réflexes simples) ou acquise (réflexes conditionnels).
I.2. Étude d'un réflexe inné (primaire, absolu ou simple)
I.2.1 Mise en évidence du réflexe inné chez la grenouille
I.2.1.1. Conditions expérimentales
L'encéphale d'une grenouille est détruit, mais la moelle épinière est laissée intacte, c'est une grenouille spinale ou médullaire. Elle est suspendue à une potence où elle reste inerte mais réagit à des excitations suffisantes (mécanique, thermique, électrique ou chimique).
I.2.1.2. Réponses à des excitations d'intensités croissantes
Trempons l'extrémité de la patte postérieure gauche de la grenouille spinale dans une solution d'acide acétique diluée de concentrations croissantes.
Résultats
Pour une concentration d'acide très faible il n'y a pas de réponse, cette excitation est infraliminaire. Une augmentation de la concentration de façon croissante entraîne d'abord une flexion du pied (réflexe localisé), puis de la patte postérieure gauche excitée (réflexe unilatéral), des deux pattes postérieures (réflexe symétrique), de toutes les pattes (réflexe irradié) et de tout le corps (réflexe généralisé).
C'est la loi de Pflüger qui dit que : la réponse musculaire obtenue suite à une excitation localisée de la peau est d'autant plus étendue que l'excitation est grande.
I.2.1.3. Structures indispensables à l'accomplissement d'un réflexe
Expériences
L'application d'éther sur la patte postérieure gauche, suivie d'une excitation supraliminaire ou suffisante, n'entraîne pas une flexion des pattes. L'éther rend insensible les récepteurs sensoriels. Donc les récepteurs sensoriels sont indispensables à la réalisation d'un mouvement réflexe.
Sectionnons le nerf sciatique ou mixte de la patte postérieure gauche et excitons la, elle ne se contracte pas. Donc le nerf est indispensable à la réalisation d'un mouvement réflexe.
La destruction de la moelle épinière suivie de l'excitation de la patte postérieure gauche n'entraîne aucune réaction. Donc l'existence d'un centre nerveux est indispensable à la réalisation d'un mouvement réflexe.
La destruction des muscles de la patte postérieure gauche suivie de son excitation n'entraîne pas sa flexion. Donc un organe effecteur (ici les muscles) est indispensable à la réalisation d'un mouvement réflexe.
Conclusion
La réalisation d'un réflexe fait intervenir :
$-\ \ $ Un récepteur sensoriel (ici les terminaisons sensorielles de la peau) ;
$-\ \ $ Un conducteur (fibres nerveuses centripètes et centrifuges) ;
$-\ \ $ Un centre nerveux (ici la moelle épinière)
$-\ \ $ Un effecteur (ici le muscle fléchisseur de la jambe).
Tout se passe comme si l'influx nerveux est réfléchi par la moelle épinière à la façon d'une lumière sur un miroir, d'où le nom de réflexe donné à cette forme d'activité nerveuse.
Le trajet suivi par l'influx nerveux depuis le récepteur jusqu'aux effecteurs est appelé arc réflexe.
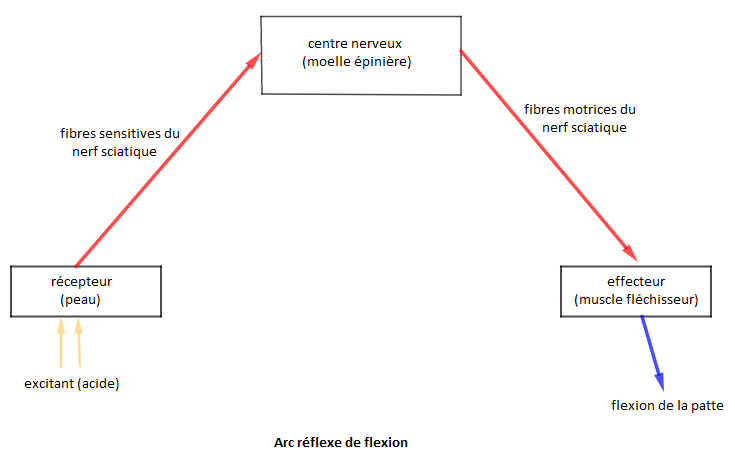
I.2.1.4. Trajet de l'influx nerveux
I.2.1.4.1. Expérience de dégénérescence de Wallérienne
La section du nerf rachidien entraîne la dégénérescence des fibres de la partie centrifuge.
La section de la racine dorsale juste avant le ganglion spinal, entraîne la dégénérescence des fibres centripètes (bout relié à la moelle épinière), mais les fibres centrifuges de cette racine et les fibres de la racine ventrale demeurent. Si la section est réalisée après le ganglion spinal, les fibres centrifuges (bout communiquant avec la patte) qui continuent dans le nerf rachidien dégénèrent.
La section de la racine ventrale entraîne la dégénérescence de ses fibres centrifuges qui continuent dans le nerf rachidien.
Conclusion
La racine dorsale contient des neurones en T dont le corps cellulaire se trouve dans le ganglion spinal, alors que la racine ventrale contient des neurones dont le corps cellulaire se trouve dans la corne ventrale de la moelle épinière et le nerf rachidien contient des fibres des racines ventrale et dorsale.
I.2.1.4.2. Expérience de Bell et Magendie
Expérience
Pour trouver le rôle joué par les racines rachidiennes dans la réalisation des réflexes de flexion Bell et Magendie réalisent les expériences suivantes: expériences de section et d'excitation.
figure....
Conclusion
Ces expériences montrent que la racine postérieure (dorsale) conduit l'influx sensitif ou centripète, tandis que la racine antérieure conduit les influx moteurs ou centrifuges et le nerf rachidien contient à la fois les fibres sensitives et motrices, c'est un nerf mixte.
I.2.1.4.3. Cas de réflexes localisé et unilatéral
Le temps parcouru par l'influx nerveux pour passer des récepteurs sensoriels aux effecteurs est plus long que celui parcouru par un influx sur une fibre nerveuse. En effet, ce temps permettrait à l'influx de traverser au moins deux synapses avant d'arriver à l'effecteur. On peut donc penser à l'existence d'un neurone appelé neurone d'association ou neurone intermédiaire ou interneurone entre le neurone sensitif et le motoneurone.
I.2.1.4.4. Cas du réflexe symétrique
La flexion de la patte opposée est due à l'existence de neurones d'association transversaux horizontaux qui transmettent l'influx nerveux du neurone sensitif au motoneurone de cette patte.
I.2.1.4.5. Cas des réflexes irradié et généralisé
Le passage de l'influx nerveux à tous les membres et muscles du corps, suppose l'existence de neurones d'association verticaux en plus des neurones d'association transversaux horizontaux transmettant l'influx aux motoneurones situés à des étages différents de la moelle épinière.
I.2.2. Caractères des réflexes innés ou simples
Un réflexe a plusieurs caractéristiques :
$-\ \ $ Il est inné c'est-à-dire acquis dès la naissance ;
$-\ \ $ Il est involontaire ;
$-\ \ $ Il est identique chez tous les êtres vivant de la même espèce (stéréotypés, spécifiques) ;
$-\ \ $ Il se produit obligatoirement (inévitable, prévisible) ;
$-\ \ $ Il est adapté à un but précis, la protection.
I.2.3. Quelques réflexes innés
Les réflexes innés sont nombreux, on peut citer quelques réflexes :
$-\ \ $ L'irritation de la cornée de l'œil par un corps étranger entraîne l'occlusion des paupières, c'est le réflexe d'occlusion des paupières.
$-\ \ $ Le chatouillement de la plante des pieds d'un sujet endormi entraine involontairement la flexion des orteils, c'est le réflexe plantaire.
$-\ \ $ Un coup sec appliqué au dessous du genou d'un sujet assis sur le bord d'une table les jambes pendantes, entraîne l'extension de la jambe, c'est le réflexe rotulien.
$-\ \ $ La percussion légère du tendon d'Achille d'un sujet assis sur le bord d'une table les jambes pendantes, entraîne l'extension du pied, c'est le réflexe achilléen.
I.2.4. Classification des réflexes
I.2.4.1. Classification suivant le centre nerveux
Suivant le centre nerveux on distingue :
$-\ \ $ Les réflexes encéphaliques dont le centre nerveux est l'encéphale : les réflexes bulbaires (salivation, respiration), les réflexes thalamiques (accommodation), les réflexes liés au cervelet (équilibration).
$-\ \ $ Les réflexes médullaires dont le centre nerveux est la moelle épinière : les réflexes de flexion.
I.2.4.2. Classification suivant la position du récepteur
Selon la position du récepteur on distingue : les réflexes extéroceptifs et intéroceptifs.
I.2.4.2.1. Les réflexes extéroceptifs
Les récepteurs sont situés à l'extérieur du corps (à la périphérie) et mettent ainsi l'organisme en relation avec son environnement. Ces récepteurs sont : tactiles, visuels, auditifs, olfactifs ou gustatifs. Ces réflexes protègent l'organisme des dangers de son milieu. Leur arc réflexe contient un neurone d'association.
I.2.4.2.2. Les réflexes intéroceptifs
Les récepteurs sont situés à l'intérieur de l'organisme et renseignent sur l'état des organes internes. On distingue : les réflexes proprioceptifs et les réflexes viscéroceptifs.
$-\ \ $ Les réflexes proprioceptifs
Ce sont des réflexes dont les récepteurs sont situés dans l'organe effecteur. C'est le cas des réflexes myotatiques (réflexes rotulien et achilléen) dont les récepteurs sont constitués par les fuseaux neuromusculaires (situés dans le muscle) et les corpuscules tendineux de Golgi (situés dans les tendons).Ces récepteurs sont sensibles à l'étirement du muscle ou des tendons d'où leur nom de mécanorécepteurs. En effet, l'étirement de ces récepteurs pendant l'allongement du muscle entraîne la naissance d'influx nerveux provoquant la contraction du muscle et son raccourcissement. Ainsi un muscle étiré tend à revenir à sa position initiale. Ces réflexes assurent l'équilibration et la coordination des mouvements.
L'arc réflexe des réflexes myotatiques ne possède pas de neurone d'association, ce sont des réflexes monosynaptiques. La réaction obtenue est plus rapide que dans un réflexe extéroceptif.
$-\ \ $ Les réflexes viscéroceptifs
Les récepteurs sont situés dans les parois des viscères (vessie, tube digestif, vaisseaux sanguins, cœur...) et captent toute variation du milieu intérieur. Ces réflexes assurent la régulation du fonctionnement des organes internes.
I.2.4.2.3. Quelques exemples de réflexes de coordination
$-\ \ $ Réponse des muscles antagonistes
La contraction d'un muscle (exemple du biceps) s'accompagne obligatoirement d'un relâchement du muscle antagoniste (le triceps). En effet, l'excitation de la main entraîne la naissance d'un influx nerveux qui emprunte les fibres sensitives, au niveau de la moelle épinière l'arrivée de l'influx nerveux excite le motoneurone innervant le biceps, alors que les neurones d'association inhibent les motoneurones du muscle antagoniste. On parle de réflexe d'innervation réciproque des deux muscles antagonistes.
$-\ \ $ Contrôle de la tension : réflexe myotatique inverse
L'étirement d'un muscle lors d'un choc au niveau de la rotule entraîne l'excitation des mécanorécepteurs. Ces derniers envoient un influx nerveux vers les motoneurones. Au niveau de la substance grise le motoneurone innervant le muscle extenseur est excité, alors que le motoneurone fléchisseur est inhibé grâce à la présence d'un interneurone. Ce qui entraine une contraction du muscle extenseur et une décontraction du muscle fléchisseur qui lui est antagoniste.
I.3. Étude de réflexe conditionnel
I.3.1. Mise en évidence de la salivation innée
Donnons à un chien muni d'une fistule salivaire des aliments (secs, solides, de la viande...) la salive coule en grande quantité. Cette salivation est due à la composition chimique et physique des aliments. Donc ces aliments sont des excitants absolus.
Cette salivation se produit même lorsque les hémisphères cérébraux sont détruits, elle est donc indépendante de la volonté et se produit avec n'importe quel chien, c'est donc un réflexe inné. Mais la section des nerfs rattachés aux glandes salivaires ou la destruction du bulbe rachidien entraîne fait disparaître ce réflexe.
La stimulation de la muqueuse buccale est un stimulus inconditionnel, car ne nécessite pas une préparation préalable pour agir.
I.3.2. Réflexe conditionnel répondant ou pavlovien
I.3.2.1. Conditions expérimentales
I.3.2.2. Réalisation du réflexe conditionnel
La réalisation de l'expérience se réalise en trois temps :
$-\ \ $ Pavlov donne des aliments au chien. Il se produit aussitôt une salivation du chien.
$-\ \ $ Quelques heures après, l'expérimentateur fait sonner le métronome. Il n'observe aucune salivation.
$-\ \ $ Quelques heures après, l'expérimentateur fait sonner le métronome et donne des aliments au chien, il salive. L'expérimentateur répète plusieurs fois cette expérience et il se produit à chaque fois une salivation. A la longue le son du métronome sans apport d'aliment fait saliver le chien. Ce résultat est un réflexe conditionnel.
I.3.2.3. Interprétation
La stimulation de la langue par les aliments entraîne une excitation du centre salivaire qui stimule, à travers les fibres sécrétrices, les glandes salivaires qui se mettent à saliver, c'est le réflexe inné de salivation.
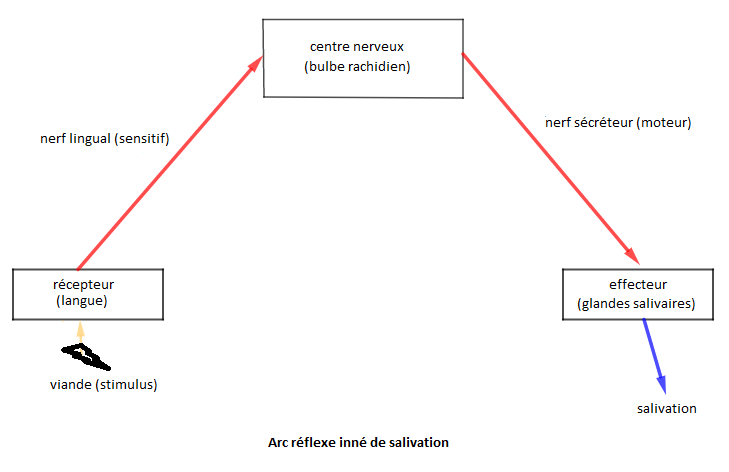
Des liaisons se créent entre les centres auditif et salivaire lors de l'association du métronome (stimulus neutre) et de l'aliment (stimulus absolu).
Le métronome seul provoque la salivation. En effet, de nouvelles liaisons se sont créées entre le centre auditif et le centre salivaire rendant le stimulus auditif efficace. Pavlov parle alors de réflexe salivaire conditionnel.
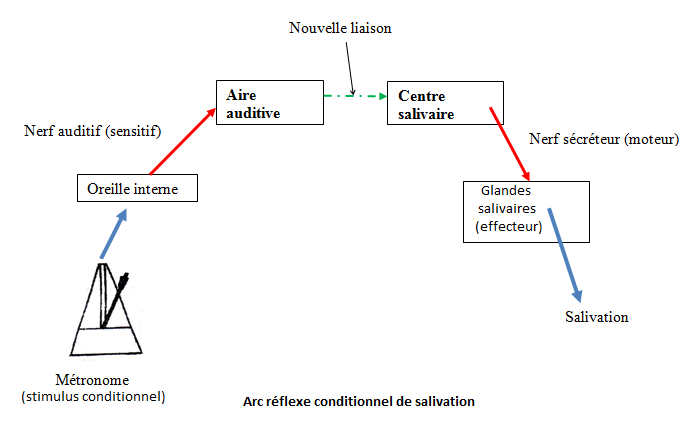
I.3.3. Les réflexes opérants ou skinnériens
I.3.3.1. Expérience de conditionnement
Skinner met un rat dans une cage riche en objets divers, dont un levier et un distributeur de nourriture qui donne de la nourriture au rat à chaque fois qu'il appuie sur un levier.
L'animal en explorant la cage appuie accidentellement sur le levier et obtient en guise de récompense de la nourriture. Le geste est accidentel la première fois, mais très vite, l'action est renouvelée et se reproduit avec une fréquence de plus en plus grande grâce à l'effet de "récompense" (renforcement) que constitue la nourriture obtenue, on dit que le renforcement est positif.
Le comportement opérant peut expliquer un grand nombre de conduites acquises au contact de l'environnement et le dressage des animaux.
Une liaison nerveuse nouvelle est formée entre l'œil qui voit le levier (le récepteur sensoriel) et les muscles moteurs de la patte qui appuie sur la pédale (l'effecteur).
Remarque
Dans d'autres circonstances, le sujet peut déclencher une punition, il apprend à éviter cette situation. On dit que le renforcement est négatif.
I.3.3.2. Comparaison entre le réflexe de Skinner et de Pavlov
Le conditionnement opérant diffère du conditionnement classique pavlovien sur des points essentiels :
$-\ \ $ Dans le conditionnement pavlovien le sujet subit le milieu, il ne le modifie pas, il s'y adapte. Il répond au stimulus conditionnel fourni par l'expérimentateur. De même, c'est l'expérimentateur qui fournit l'excitant absolu pendant le conditionnement.
$-\ \ $ Dans le conditionnement opérant, c'est le sujet lui-même qui agit sur le milieu accidentellement d'abord, puis de façon automatique ou volontaire. C'est le sujet qui déclenche l'apparition de la récompense.
I.3.4. Les caractères du réflexe conditionnel
Les réflexes conditionnels présentent plusieurs caractéristiques :
$-\ \ $ Ils sont acquis, dépendent de l'expérience personnelle d'un individu ;
$-\ \ $ Ils sont temporaires et non immuables, en effet, non entretenus, ils disparaissent ;
$-\ \ $ Ils sont obtenus par l'association de n'importe quelles excitants, l'essentiel est que l'excitant conditionnel soit précisé et qu'il précède qu'il précède toujours l'excitant absolu (en effet, le stimulus conditionnel doit être bien déterminé, car des stimuli divers peuvent déclencher ce réflexe, ainsi la vue ou l'odeur d'un aliment peut devenir un excitant conditionnel) ;
$-\ \ $ Ils exigent l'intervention du cerveau au niveau duquel s'établissent les nouvelles liaisons.
I.3.5. Importance du réflexe conditionnel
Le réflexe conditionnel est très important dans la vie d'un être vivant, en effet il intervient lors de :
$-\ \ $ L'habituation (accoutumance) qui est la première manifestation qui apparaît chez le nourrisson.
$-\ \ $ L'apprentissage associatif qui est le résultat de plusieurs conditionnements.
$-\ \ $ Le dressage des animaux pour le spectacle (lions, singes...), pour la recherche de drogues, de bombes (chiens des policiers). L'animal apprend par l'entrainement à bien réagir à des signaux.
$-\ \ $ Chez l'homme l'apprentissage permet :
$\ast\ \ $ l'acquisition d'automatismes (conduite de voiture, équilibre sur le vélo...) ;
$\ast\ \ $ la mise en place de moyens de communication entre individus (parole, écriture, lecture...) ;
$\ast\ \ $ l'élaboration de signaux intervenant dans les rapports sociaux, l'éducation (saluer...) ;
$\ast\ \ $ l'adaptation individuelle.
I.3.6. Comparaison entre les réflexes innés et conditionnels
$$\begin{array}{|c|c|c|}\hline\text{Réflexes}&\text{Inné}&\text{Conditionnels ou acquis}\\ \hline\text{Ressemblances}&\text{Réponses involontaires}&\text{Réponses involontaires}\\ \hline&\text{Inné}&\text{Acquis par l'apprentissage}\\&\text{Héréditaires}&\text{Individuels}\\ \text{Différences}&\text{Spécifiques}&\text{Disparaît sans entretien}\\&\text{Absolus}&\text{Conditionnels}\\& \text{Immuables}&\\ \hline\end{array}$$
Conclusion
Les réflexes sont nombreux et sont innés ou conditionnels. Ils présentent des caractéristiques qui leur sont propres et jouent un rôle important dans la survie de l'être vivant et ont des centres nerveux divers.
II. L'activité volontaire
II.1. Notion de mouvement volontaire
Observations
$-\ \ $ Un élève décide de noter par écrit une idée, il prend son stylo et écrit
$-\ \ $ Un promeneur marche dans la rue et fait face à un obstacle et décide de le contourner par la gauche
Interprétation
Dans la première situation l'acte d'écrire est précédé d'une intention.
Dans la seconde, l'individu reconnait un obstacle. C'est-à-dire un facteur qui a modifié ses rapports avec l'environnement. Il prend conscience de sa nouvelle situation et choisit de contourner l'obstacle. Ce choix fait intervenir la mémoire et les facultés intellectuelles donc une prise de décision.
Dans tous les deux cas la décision aboutit à une réaction motrice bien adaptée. On parle de motricité volontaire ou dirigée.
II.2. Importance du cerveau dans l'activité volontaire
II.2.1. Observation
$-\ \ $ Un nouveau né privé d'hémisphères cérébraux (mal formation congénitale) est aveugle, ne manifeste aucune émotion et ne sort jamais de sa léthargie (paresse, inerte).
$-\ \ $ Des anesthésiques généraux atteignent le cerveau et suppriment toute conscience chez l'homme.
$-\ \ $ Des animaux qui ont subi l'ablation du cortex cérébral deviennent des automates.
Conclusion
Le cerveau intervient dans les comportements sensori-moteurs.
Expériences
$-\ \ $ L'ablation d'un hémisphère cérébrale chez l'homme provoque la perte de la motricité volontaire de la moitié du corps opposée à l'hémisphère cérébrale enlevé. On parle d'hémiplégie
$-\ \ $ Chez l'animal, l'ablation d'un territoire du cerveau situé en avant du sillon de Rollando provoque une paralysie musculaire de la partie du corps opposée à cette hémisphère.
$-\ \ $ Si cette ablation porte sur une petite surface, on a diminution de la partie corporelle paralysée.
Conclusion
Donc on a une subdivision du cerveau en territoires spécialisés. On parle de localisation cérébrale.
Des excitations portées sur des territoires précis des hémisphères cérébraux ont mis en évidence l'existence de relation entre des zones corticales bien localisées et des régions bien définies du corps. Le cerveau est donc responsable de la motricité dirigée grâce à la localisation des territoires sensori-moteurs spécialisés.
II.3. Les localisations cérébrales
II.3.1. Technique d'investigation
Les connaissances sur la motricité dirigée repose sur les faits d'observation, des expériences d'ablation totale ou partielle, des expériences d'excitation.
Mais en plus de ces méthodes on a les techniques suivantes :
$-\ \ $ L'électroencéphalographie cérébrale :
On enregistre l'activité électrique du cerveau grâce à des électrodes placées sur le cuir chevelu.
Les tracés obtenus appelés électroencéphalogramme montrent des oscillations appelées ondes cérébrales. Ces ondes sont variable selon l'état physiologique de l'individu.
$\ast\ \ $ Les ondes $\alpha$ sont des oscillations régulières de grande amplitude mais de fréquence faible. Elles caractérisent un état de repos mental quand les yeux sont fermés.
$\ast\ \ $ Les ondes $\beta$ sont de faible amplitude mais de fréquence élevée. Elles caractérisent un état d'éveil.
$-\ \ $ La scintigraphie :
On injecte des substances radioactives ayant une affinité avec les cellules du cerveau et on suit leur distribution qui sera plus importante au niveau des cellules en activité.
$-\ \ $ La tomodensitométrie ou scannographie :
C'est une exploration du cerveau au scanner
$-\ \ $ L'angiographie :
C'est la radiographie des vaisseaux sanguins. Elle consiste à injecter dans la circulation cérébrale un produit radio opaque et on suit sa progression par une série de clichés.
$-\ \ $ L'imagerie par résonance magnétique (IRM) :
Il s'agit de placer un individu dans un champ magnétique et d'enregistrer les signaux émis par les atomes à l'aide d'un ordinateur.
Ces différentes techniques ont permis de réaliser une cartographie cérébrale.
II.3.2. La cartographie cérébrale
Elle montre une subdivision du cortex cérébrale en aires :
$-\ \ $ l'aire de projection motrice ou aire motrice principale :
Elle est située en avant du sillon de Rolando. Sa destruction dans l'hémisphère cérébrale gauche entraîne la suppression des mouvements volontaires dans la partie droite du corps et inversement.
En effet, chaque partie du corps capable de mouvement est représentée sur l'aire motrice par une surface dont l'étendu ne dépend pas du volume des muscles mis en jeu mais est plutôt proportionnelle à la précision, la finesse et la variété des mouvements effectués. C'est pourquoi la surface commandant les mouvements de la main est beaucoup plus importante que celle qui commande les mouvement du tronc. La répartition de ces aires qui représente les différentes parties du corps correspond schématiquement à un sujet couché avec une grosse tête, une main énorme avec le pouce prépondérant, un tronc réduit et des pieds importants. Cette représentation est appelée homuculus moteur. Cette même démarche a permis de réaliser un homuculus sensitif sur l'aire de la sensibilité générale.
$-\ \ $ L'aire psychomotrice :
Elle est située en avant de l'aire de projection motrice. Sa destruction est accompagnée de l'impossibilité à accomplir des gestes après, alors que les muscle ne sont pas paralysés ; on parle d'apraxie. Cette aire joue un rôle de coordination.
$-\ \ $ l'aire somesthésique (ou aire de la sensibilité générale) :
Elle se situe en arrière du sillon de Rolando et reçoit les messages provenant de la sensibilité générale. A côté de cette aire on a une aire psychosensorielle permettant la perception des sensations.
$-\ \ $ les aires sensorielles spécialisées :
Ce sont les aires reliées aux organes des sens : aire visuelle, aire auditive, aire gustative et aire olfactive.
II.4. Trajet des influx nerveux dans les mouvements volontaires
II.4.1. Les aires motrices
Elles sont constituées de deux types de voies.
a) Les voies directes ou pyramidales
Ce sont des neurones qui partent du cortex cérébrale, croisent le plan de symétrie au niveau du bulbe rachidien ou de la moelle épinière dans laquelle ils entrent en contact synaptique avec les moto-neurones innervant les muscles du côté opposé de l'hémisphère cérébrale correspondant. Ces voies sont mono-synaptiques et permettent les mouvement fins et précis.
b) Les voies indirectes ou extra pyramidales
Elles quittent le cortex pré-moteur et établissent des synapses au niveau des centres sous-corticaux et dans les noyaux gris du tronc cérébrale. Ces voies sont polysynaptiques et moins rapides. Elle interviennent dans la commande des mouvements d'ensemble et dans le contrôle des postures.
II.4.2. Les voies sensorielles
Ce sont des fibres nerveuses en provenance de divers récepteurs sensoriels (organes des sens, récepteurs musculaires, tendineux et articulaires). Elles remontent la moelle épinière dans sa région dorsale et après des relais synaptiques atteignent l'aire sensoriel du cortex de l'hémisphère cérébrale opposé. Ce sont les voies ascendantes de la sensibilité consciente.
Auteur:
Mamadou Ba
Commentaires
Anonyme (non vérifié)
lun, 12/16/2019 - 02:35
Permalien
Intéressant
Anta (non vérifié)
dim, 11/08/2020 - 03:50
Permalien
Vous nous êtes d’une très
Abou Moussa Sarr (non vérifié)
mar, 04/05/2022 - 23:43
Permalien
Travail
Abou Moussa Sarr (non vérifié)
mar, 04/05/2022 - 23:43
Permalien
Travail
ibrahima sao (non vérifié)
jeu, 06/02/2022 - 00:29
Permalien
Excellent travail
Ajouter un commentaire